

1. 서 론
2. 재료 및 방법
2.1 조류 배양
2.2 Nq 4-6의 살조 범위 탐색
2.3 Nq 4-6의 미소생태 실험
2.4 Nq 4-6에 의한 식물플랑크톤 군집 구조 분석
3. 결과 및 고찰
3.1 Nq 4-6의 살조 범위 탐색
3.2 Nq 4-6에 의한 수환경 및 군집 구조 변화
4. 결 론
1. 서 론
우리나라는 겨울철 저수온기에 매년 한강 및 낙동강에서 규조 Stephanodiscus hantzschii에 의한 대발생이 빈번하게 발생하고 있다 (Cho et al. 1998, Ha et al. 2003, Jung et al. 2008, Kang et al. 2011). S. hantzschii를 포함한 규조류 대발생은 수계의 심미적 가치 하락, 이취미로 인한 식수원 확보 문제 및 수면 휴양 시설의 가치하락 등의 문제를 일으키며, 특히 정수처리시설에 문제를 일으켜 경제적 손실을 발생시킨다 (Bourne et al. 1996, Dawson 1998, Choi et al. 2005). 또한 수계 내 용존산소의 고갈 및 수중 생물의 사멸 등 수중 생태계에 심각한 영향을 초래한다 (Lee et al. 2001, Han et al. 2002).
현재까지 조류제어를 위해 다양한 방법 등이 개발되어 사용되고 있으나 일부 방법은 난분해성 물질을 이분해성으로 전환시켜 조류의 대발생을 더욱 촉진시킬 가능성을 내포하고 있으며, 수중 및 저니토에 축적되어 생태독성의 잠재적인 위험성을 갖고 있다 (Park et al. 2001). 또한 현재 국내·외에서 주로 사용되는 황토, Phoslock® 등의 응집, 탈인제 등의 조류 제어 기술은 대발생 원인 종 외에 다른 생물에 영향을 미쳐 수중 생태계가 파괴될 수 있다 (Lim et al. 2002, Yamamoto et al. 2005, Lürling and Oosterhout 2013). 이와 같은 무분별한 조류 제어는 생물 종 다양성 감소 및 먹이사슬 교란 등의 수중생태계 파괴로 야기될 수 있다. 최근 세계적으로 생물 종 다양성에 대한 중요성이 부각되고 있는 실정이며, 국내 역시 수질개선에 대한 개념이 물리화학적 수질개선에서 생태적으로 건강한 수질개선으로 변화하고 있는 추세이다 (Kim et al. 2008, Hong 2014). 따라서, 조류 제어를 위해 대상 조류에 특이적인 살조범위를 갖고 생태계에 교란이 적은 친환경적인 조류 제어 기술 개발의 필요성이 대두되고 있다 (Gumbo et al. 2008, Byun et al. 2014).
본 연구진은 보다 친환경적인 조류 제어 기법을 개발하고자 자연계에 존재하는 물질을 대상으로 탐색하였고, 그 결과 Naphthoquinone (이하 Nq)을 기반으로 한 생물유래 합성물질을 개발하였다 (Joo 2012). Nq 유도체는 다양한 식물과 미생물에서 생산되는 물질로 항암제, 항증식제, 항혈소판제, 항염증제, 항알레르기제 등의 기능뿐만 아니라, 다양한 세균, 곰팡이 등에도 효과적인 제어효과를 나타내는 물질로 의약품에 많이 쓰인다 (O’Brien 1991, Monks et al. 1992, Dong et al. 2009, Menna-Barreto et al. 2009). 합성된 물질 중 겨울철 대발생을 일으키는 규조 S. hantzschii에 가장 살조 활성이 높았던 Nq 4-6 화합물을 선별하였다. 하지만 최종 선정된 Nq 4-6화합물이 S. hantzschii를 제외한 다른 조류에 대한 영향을 알아보는 실험은 진행되지 않았으며, 선정된 화합물을 현장수 적용 시 발생하는 미소생태 변화에 대해 연구된 바 없었다.
따라서 본 연구에서는 S. hantzschii에 대해 높은 살조 활성을 나타냈던 Nq 4-6 화합물의 살조 범위에 대해 실험을 진행하였다. 또한, Nq 4-6 화합물의 적용 가능성을 평가하기 위해 현장수를 이용한 미소생태 실험으로 Nq 4-6 화합물 적용 후 물리화학적 및 생물학적 요인 변화를 관찰하였다.
2. 재료 및 방법
2.1 조류 배양
본 연구에 사용된 식물플랑크톤은 총 12종으로 남조 Aphanizomenon flos-aquae (HYL1112-AP1), Dolichospermum circinale (HYCC201307-AN13), Microcystis aeruginosa (HYK0906-A2), Oscillatoria sp. (HYHC1409-OS01) 4종, 규조 Aulacoseira gra-nulata (HYND1404AGZ2), Cyclotella meneghiniana (HYND1404CMZ3), Fragilaria acus (HYND1404 SAZ1)와 Stephanodiscus hantzschii (HYHC1405- SH01) 4종은 모두 본 연구진이 북한강 및 낙동강에서 출현한 종을 직접 분리 및 배양하였다. 남조 Nostoc paludosum (KMMCC-1184), 녹조 Scenedesmus qua-dricauda (AG10003) 2종은 한국 생명공학연구원 (KRIBB)에서 분양받았으며, 녹조 Cosmarium bio-culatum (CCAP 612/17) 1종은 영국 조류 및 원생동물 배양 센터 (CCAP, Culture Collection of Algae and Protozoa, SAMS Research Service Ltd, Scottish Marine Institute, England)에서 각각 분양받았다 (Table 1).
남조류 및 녹조류의 경우 CB 배지 (NIES, National Institute for Environmental Studies, Japan), 20°C, 광도 30-40 μmol m-2 s-1, 광주기 12 : 12 (dark : light), 규조류의 경우 15°C, DM media (CCAP, Culture Collection of Algae and Protozoa, SAMS Research Service Ltd, Scottish Marine Institute, England), 15°C, 광도 30-40 μmol m-2 s-1, 광주기 12 : 12 (dark : light)의 조건으로 배양하였다.
2.2 Nq 4-6의 살조 범위 탐색
Naphthoquinone Nq 4-6 화합물이 규조 Stephano-discus hantzschii를 포함한 다른 조류에 어떤 영향을 미치는지 평가하기 위해 남조류, 녹조류 및 규조류에 Nq 4-6 화합물을 최종농도 0.5, 1, 5 및 10 μM로 접종하여 살조 활성을 관찰하였다. 남조 Aphanizomenon flos-aquae (HYL1112-AP1), Dolichospermum cir-cinale (HYCC201307-AN13), Microcystis aeruginosa (HYK0906-A2), Nostoc paludosum (KMMCC-1184) Oscillatoria sp. (HYHC1409-OS01) 녹조 Scenedesmus quadricauda (AG10003), Cosmarium bioculatum (CCAP 612/17) 및 규조 Aulacoseira granulata (HYND1404AGZ2), Cyclotella meneghiniana (HY ND1404CMZ3), Fragilaria acus (HYND1404SAZ1), Stephanodiscus hantzschii (HYHC1405-SH01)를 각각 5 x 103 cell mL-1 밀도로 48 well plate (1 mL)에서 실험을 수행하였다. 살조 활성은 Nq 4-6 화합물을 접종 후 7일째 계수한 결과를 바탕으로 다음과 같은 식으로 계산하였다.
살조 활성 (algicidal activity, %) = 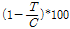 (1)
(1)
상기 식에서 C: 물질을 접종하지 않은 대조구의 세포 수와 T: 물질 접종 7일 후 세포 수를 의미한다.
Nq 4-6 화합물에 대하여 각 조류 별 살조 활성 (%) 결과를 기반으로 하여 50%의 살조활성을 나타내는 effect concentration (EC50)값을 도출하였다. EC50 값은 다음과 같은 식으로 Sigmaplot Software (USA)를 이용하여 측정하였다 (Kooijman 1987).
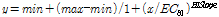 (2)
(2)
2.3 Nq 4-6의 미소생태 실험
Nq 4-6 화합물의 현장적용 가능성을 평가하기 위해 매년 규조 S. hantzschii에 의해 조류 대발생에 의해 피해가 심각한 낙동강 현장수를 채수하여 미소생태 실험을 수행하였다. 미소생태 실험은 2014년 2월 낙동강 삼랑진 (35o 22′ 28˝ N, 128o 38′ 23˝ E)에서 채수하여 냉암소 상태로 실험실로 운반 후 수행하였다. 채수한 현장수는 S. hantzschii가 8 x 103 cells mL-1 밀도로 우점하였으며, 실험은 10°C, 광도 30-40 μmol m-2 s-1, 12 : 12 cycle (dark : light)의 조건에서 10 L 규모로 진행하였다. Nq 4-6 화합물에 대한 미소생태 실험은 선행연구를 통해 효과가 입증된 농도인 0.5 μM을 최종농도로 접종하여 15일간 진행되었으며, 각각의 실험구는 S. hantzschii가 침전되는 것을 방지하기 위해 수중펌프를 이용해 물을 혼합시켜 주었다.
물리화학적 요인 중 수온, pH, 전기전도도, 탁도는 portable multiparameter (HORIBA U50, Japan)로 직접 측정하였고 용존산소 (DO), 생화학적 산소요구량 (BOD5), 영양염 항목은 채수 후 측정하였다 (APHA 2005). 용존산소는 Winkler 법에 따라 측정하였으며, 생화학적 산소요구량도 Winkler 법에 따라 측정하였다. 암모니아성 질소 (NH4-N), 아질산성 질소 (NO2-N), 질산성 질소 (NO3-N), 인산염 인 (PO4-P), 총 인 (TP) 및 규산염 규산 (SiO2-Si)은 APHA (2005)을 따랐다.
식물플랑크톤 변화는 선정된 Nq 4-6의 처리 당일을 0일로, 처리 후 2, 4, 7, 10, 15 일째 마다 50 mL를 분취하여 Lugol 용액으로 고정한 뒤 세포를 계수하였다. 식물플랑크톤의 동정은 광학 현미경 (Olympus IX71, Japan)의 x 200~400배율에서 Sedgwick-Rafter Chamber (PhytoTech Inc., USA)를 이용하여 종 수준까지 동정하였고, 동일 속에서 뚜렷한 식별 형질의 차이를 보이지 않는 종은 미동정종 (sp.)으로 처리하였다. 세균 및 heterotrophic nanoflagellates (HNFs)의 경우, 각각 2 mL, 10 mL을 분취하여 DAPI (4’,6’- diamidino-2-phenylindole) 법으로 5분간 염색한 후, 0.2 μm-pore-size black polycarbonate filters (Millipore Inc., USA)로 포집한 후 x1000 배율의 형광 현미경 (Olympus BX51, Japan)로 발광하는 세포를 계수하였다 (Porter and Feig 1980). HNFs 세포는 red chlorophyll fluorescence가 관찰되는 세포를 계수하였다 (Kimio et al. 1996). Ciliate는 50 mL을 분취하여 Lugol 용액으로 고정한 뒤 식물플랑크톤과 동일한 방법으로 세포를 계수하였다.
2.4 Nq 4-6에 의한 식물플랑크톤 군집 구조 분석
Nq 4-6 화합물에 의한 식물플랑크톤 군집의 변화를 알아보기 위해 우점도 지수 (dominance index), 다양도 지수 (diversity index), 풍부도 지수 (richness index), 균등도 지수 (evenness index) 등을 Nq 4-6 화합물을 적용하지 않은 대조구와 Nq 4-6 화합물을 적용한 처리구에서 각각 산출하였다.
우점도 지수 (dominant index)의 경우 우점종과 아우점종을 선정하여 계산하였으며, 지수의 산출은 McNaughton의 우점도 지수에 의하여 산출하였다 (McNaughton 1967).
DI = (n1+ n2) (3)
상기 식에서 N: 총 현존량, n1: 우점종 현존량 및 n2: 아우점종 현존량을 의미한다.
다양도 지수의 경우 Margalef (1958)의 정보이론에 의해 유도된 Shannon-Wiever의 식 (Pielou 1969)을 이용하여 산출하였다.
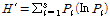 (4)
(4)
상기 식에서 H': 다양도, S: 전체 종수 및 Pi: i 번째에 속하는 개체수의 비율을 말하며 (ni/N)으로 계산 (N: 군집내의 전체 현존량, ni: 각 종의 현존량)하는 것을 의미한다.
풍부도 지수의 경우 총 현존량과 종수 만을 가지고 군집의 상태를 표현하는 지수로서 Margalef (1958)의 지수를 사용하여 산출하였다.
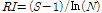 (5)
(5)
상기 식에서 RI: 풍부도, S: 전체 종수, N: 전체 현존량을 의미한다.
균등도 지수의 경우 각 지수의 최대치에 대한 실제치의 비로써 표현된다. 각 다양도 지수는 군집내 모든 종의 개체수가 동일할 때 최대가 되므로 결국 균등도 지수는 군집내 종 구성의 균일한 정도를 나타내는 것으로 Pielou (1975)의 식을 사용하여 산출하였다.
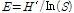 (6)
(6)
상기 식에서 E: 균등도, H': 다양도, S: 전체 종수를 의미한다.
3. 결과 및 고찰
3.1 Nq 4-6의 살조 범위 탐색
Naphthoquinone Nq 4-6 화합물이 Stephanodiscus hantzschii를 포함한 다양한 조류에 미치는 영향을 평가하기 위하여 남조류 5종, 녹조류 2종, 규조류 4종에 접종 후 7일째까지 50 %의 살조 활성을 나타내는 EC50 값을 도출하였다 (Fig. 1, Table 2). Nq 4-6 화합물 접종 7일째 남조 A. flos-aquae (HYL1112- AP1)의 EC50 값은 9.4 μM로 나타났고, 남조 D. circinale (HYCC201307-AN13)의 EC50 값은 5.3 μM로 나타났다. 남조 M. aeruginosa (HYK0906-A2)의 EC50 값은 8.2 μM로 나타났고, 남조 N. paludosum (KMMCC-1184)의 EC50 값은 6.5 μM로 나타났으며, 남조 Oscillatoria sp. (HYHC1409-OS01)의 EC50 값은 5.3 μM로 나타났다. 녹조 S. quadricauda (AG 10003)의 경우 접종 최고농도인 10 μM에서 50% 미만의 살조 활성으로 EC50 값이 나타나지 않았으며, 녹조 C. bioculatum (CCAP 612/17)의 EC50 값은 4.7 μM로 나타났다. 또한 규조 A. granulata (HYND 1404AGZ2)의 EC50 값은 0.9 μM로 나타났고, 규조 F. acus (HYND1404SAZ1)의 EC50 값은 1.7 μM로 나타났다. 규조 C. meneghiniana (HYND1404CMZ3)와 S. hantzschii (HYHC1405-SH01)의 경우 접종 최소농도인 0.5 μM에서 50% 이상의 살조 활성이 나타나 EC50 값이 도출되지 않았다.
최근까지 개발된 대부분의 조류 제어제는 비특이적인 넓은 살조 범위로 인해 수중 생태계를 파괴할 수 있는 단점이 있다 (Koss and Snyder 2005, Gumbo et al. 2010). Nq 4-6 화합물에 대한 EC50 값을 도출한 결과 남조류 및 녹조류에서 EC50 값이 모두 4 μM 이상의 농도로 나타났다. 규조류의 경우 남조류 및 녹조류 보다 낮은 EC50 값을 나타냈으며, 특히 원반형 규조류 (centric diatom)인 C. meneghiniana, S. hantzschii에서 EC50 값이 0.5 μM 이하의 농도로 도출된 것으로 미루어 볼때, 0.5 μM의 농도를 적용한다면 다른 조류에 영향을 미치지 않고, 원반형 규조류만을 특이적으로 제어할 수 있을 것으로 사료된다.
3.2 Nq 4-6에 의한 수환경 및 군집 구조 변화
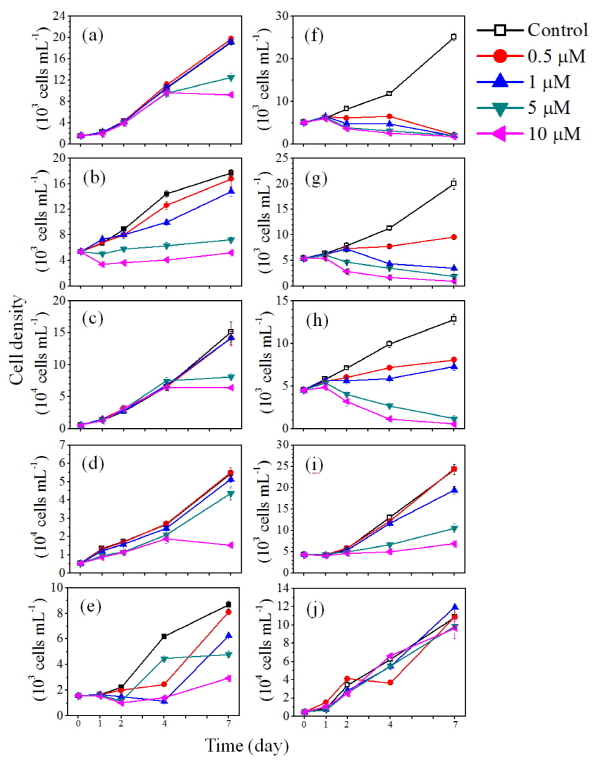
Nq 4-6 화합물의 현장적용 가능성을 평가하기 위해 규조 Stephanodiscus hantzschii 가 우점하는 낙동강 현장수를 이용한 10 L 규모의 미소생태 실험을 수행하여 물리화학적 요인 및 생물학적 요인에 대해 분석하였다. 미소생태 실험의 물리화학적 요인을 분석한 결과 수온, pH, 전기전도도, 생화학적 산소요구량 (BOD5)은 대조구와 Nq 4-6 처리구에서 실험 종료 시까지 유사한 경향성을 보였다 (Fig. 2). 탁도의 경우 대조구에서 실험 시작 시부터 10일째까지 지속적으로 증가하였다. 반면 Nq 4-6 처리구에서는 실험 시작 시부터 15일 실험 종료 시까지 지속적으로 감소하는 경향을 보였다. 이는 처리구에서 Nq 4-6 화합물에 의해 S. hantzschii 현존량이 급감 하면서 낮은 탁도 값이 측정된 반면, 대조구에서는 지속적인 세포의 증가에 의하여 높은 탁도 값이 측정된 것으로 사료된다. 용존산소 (DO)는 대조구에서 4일째까지 증가하였으나, 이후 실험 종료 시인 15일째까지 지속적으로 감소하였다. Nq 4-6 처리구에서는 실험시작 시부터 10일째까지 지속적으로 감소하였으나, 실험 종료 시인 15일째에 9.2 mg L-1로 증가하였다. 용존산소 (DO)의 경우 실험 초기에 대조구에서 증가하는 경향이 관찰되었는데, 이는 식물플랑크톤 현존량이 높아짐에 따라 광합성에 의한 산소 발생량이 높아져 증가한 것으로 판단된다 (Jacob 1960, Choi et al. 1997).
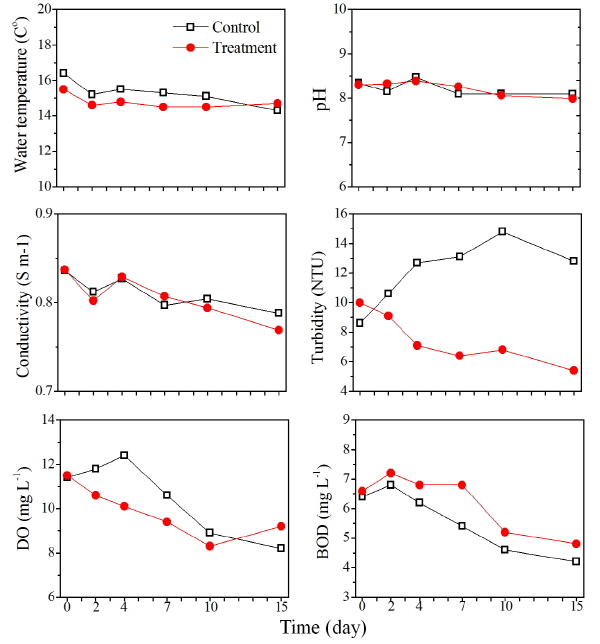
영양염 분석한 결과 총인 (TP)은 대조구에서 실험 시작 시부터 10일째까지 지속적으로 감소하였으며, 실험 종료 시인 15일째에 증가하였다 (Fig. 3). Nq 4-6 처리구에서는 실험 시작 시부터 지속적으로 감소하여 15일째 실험 종료 시까지 감소하였는데, 대조구의 경우 7일 이후 식물플랑크톤이 사멸하면서 총인이 증가한 것으로 사료되며, 처리구의 경우 실험 초기에 처리구에서 Nq 4-6에 의해 우점종인 규조 S. hantzschii가 사멸하면서 증가한 것으로 사료된다 (Proctor and Fuhrman 1990, Shigui 1993). 인산염 인 (PO4-P)은 대조구에서 실험 시작 시부터 15일째 실험 종료 시까지 지속적으로 감소하였으며, Nq 4-6 처리구에서는 실험 시작 시부터 7일째까지 감소하였으며, 10일째에 증가하였고, 실험 종료 시까지 농도가 유지되었다. 이와 같은 결과는 식물플랑크톤 현존량이 높았던 대조구에서 식물플랑크톤이 성장을 위해 인산염 인 (PO4-P)을 지속적으로 소비하여 감소한 것으로 판단된다 (Tilman and Kilham 1976, Kim et al. 2004). 아질산성 질소 (NO2-N)는 대조구와 Nq 4-6 처리구에서 모두 지속적으로 증가하는 경향을 보였으나, 대조구에서 2일째 0.21 mg L-1에서 4일째 0.79 mg L-1로 큰 폭으로 증가한 반면, 처리구에서는 비교적 낮은 양이 증가했다. 질산성 질소 (NO3-N)의 경우 대조구에서 실험기간 동안 증가, 감소가 반복되어 나타났으며, 실험 종료 시인 15일째에 0.24 mg L-1까지 감소하였다. Nq 4-6 화합물을 0.5 μM로 접종한 처리구에서는 4일째 이후부터 실험 종료 시인 15일째까지 지속적으로 감소하였다. 대조구의 경우 아질산성 질소와 질산성 질소가 4일째에 큰 폭으로 변화를 일으켰는데, 이는 아질산성 질소의 증가한 양과 질산성 질소가 감소한 양이 유사한 것으로 나타나 환원으로 인한 증가 및 감소로 사료된다. 아질산성 질소의 경우 처리구에서 대조구에 비해 낮은 농도로 측정되었다. 이는 naphthalene의 경우 아질산성 질소에 결합하여 N2로 변환시키는 작용을 하는데, naphthalene과 구조적으로 유사한 Nq 4-6 화합물이 아질산성 질소와 결합으로 인해 농도가 감소했을 것으로 사료된다 (Atkinson et al. 1990, Sasaki et al. 1997). 암모니아성 질소 (NH4-N)는 대조구와 Nq 4-6 처리구에서 모두 실험기간 동안 증가, 감소가 반복되어 나타났으며, 대조구와 Nq 4-6 처리구가 유사한 경향을 나타났다. 규산염 규소 (SiO2-Si)는 대조구에서 실험 시작 시부터 지속적으로 감소하여 10일째부터 규산염 규소가 검출되지 않았는데, 이는 대조구에서 우점했던 규조 S. hantzschii가 지속적으로 소비하여 감소한 것으로 판단된다 (Tilman and Kilham 1976, Kim et al. 2004). Nq 4-6 처리구에서는 실험 시작 시 7일째를 제외하고, 실험 종료 시인 15일째까지 지속적으로 증가하였는데, 이는 Nq 4-6 화합물로 인해 규조 S. hantzschii가 사멸하면서 규소가 용출 되었을 것으로 사료되며, 소비 또한 되지 않았을 것으로 판단된다 (Froelich et al. 1992). 전체적으로 Nq 4-6 화합물 적용 후 높은 효율로 우점종인 규조 S. hantzschii의 사멸에도 불구 하고 영양염이 크게 증가하지 않은 것으로 보아 Nq 4-6 화합물에 의한 2차적 문제로 부영양화 현상은 일어나지 않을 것으로 사료된다.
|
Fig. 2. Physicochemical parameters of water temperature, pH, turbidity, conductivity, dissolved oxygen (DO), biochemical oxygen demand (BOD) in control and treatment of naphthoquinone 4-6. |
|
Fig. 3. Nutrients variation PO4-P, TP, NO3-N, NO2-N, NH4-N and SiO2-Si in control and treatment of naphthoquinone 4-6. |
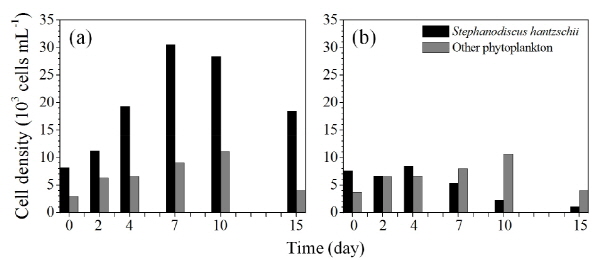
미소생태 실험의 식물플랑크톤 분석 결과 대조구에서 규조 S. hantzschii의 현존량은 실험 시작 시 8.2 x 103 cells mL-1 에서 7일째 3.1 x 104 cells mL-1까지 증가하였고, 10일째부터 감소하기 시작하여 15일째 1.8 x 104 cells mL-1까지 감소하였으나, 실험기간 동안 지속적으로 우점하였다 (Fig. 4). 7일째 이후 대조구에서 식물플랑크톤 현존량 감소는 규산염 규소가 고갈됨에 따라 우점종인 규조 S. hantzschii 및 다른 규조류가 증가하지 못해 감소한 것으로 사료되며, 규산염 규소를 이용하지 않는 녹조류의 경우 10일째까지 현존량이 증가하였다. Nq 4-6 처리구에서 S. hantzschii의 현존량은 실험시작 시 7.6 x 103 cells mL-1에서 실험기간 동안 지속적으로 감소하여, 15일째 1.0 x 103 cells mL-1까지 감소하였고, S. hantzschii에 대한 살조 활성은 현존량이 가장 높았던 7일째에 대조구 대비 85.6%의 살조활성을 나타냈다. 반면, 일부 규조류를 제외한 다른 조류의 경우 살조활성이 나타나지 않았다. 이는 Nq 4-6 화합물의 살조범위에 대한 실험 결과와 동일한 결과로 미소생태 실험에서 역시 규조류에 대해 특이적으로 높은 살조활성을 보였다.
|
Fig. 4. Changes in phytoplankton populations as a result of the algicidal effect of naphthoquinone 4-6 compound in the microcosm test. Control (a), and treatment (b) of naphthoquinone 4-6. |
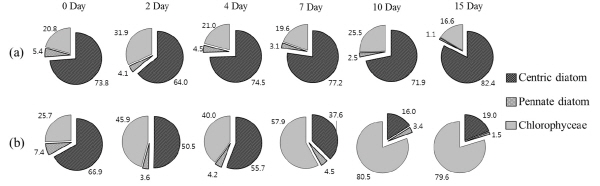
식물플랑크톤 군집의 경우 대조구에서 규조 S. hantzschii가 속한 원반형 규조류 (centric diatom)가 실험 시작 시에 전체 식물플랑크톤 대비 73.8% 우점율로 우점하였으며, 실험이 종료되었던 15일째까지 82.4%의 높은 우점율을 보였으나, 반면 Nq 4-6 처리구에서는 실험 시작 시 원반형 규조류가 66.8%의 우점율로 나타났으나, Nq 4-6 화합물에 의한 지속적인 세포수 감소로10일째에 16.0%까지 우점율이 감소하였다 (Fig. 5). 식물플랑크톤의 종조성은 대조구에서는 실험기간 동안 모두 규조 S. hantzschii가 지속적으로 우점종으로 출현하였다. 아우점종은 실험 시작부터 종료까지 녹조 Scenedesmus quadricauda가 출현하였으며, 2일째에만 녹조 Pediastrum duplex가 아우점종으로 출현하였다. Nq 4-6 처리구의 경우 실험 시작부터 7일째까지 규조 S. hantzschii가 우점종으로 출현하였고, 녹조 Scenedesmus quadricauda, Pediastrum duplex가 아우점종으로 출현하였으나, 10일째 이후부터 15일째 실험 종료 시까지 우점종이 녹조 Scene-desmus quadricauda, 아우점종은 규조 S. hantzschii로 우점종이 천이되었다. 이러한 결과는 Nq 4-6 화합물이 우점종인 규조 S. hantzschii를 제어 함으로써 종간 경쟁에서 녹조 Scenedesmus qua-dricauda가 우위를 차지해 우점하였을 것으로 사료된다 (Morin 2011, Joo 2012).
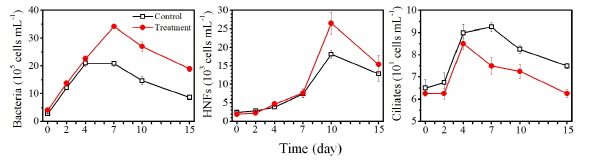
다른 생물학적 요인으로 세균을 분석한 결과, 대조구에서 실험 시작 시 1.6 x 105 cells mL-1 에서 4일째 2.1 x 106 cells mL-1까지 증가하였고, 이후 실험 종료 시 15일째 8.6 x 105 cells mL-1까지 지속적으로 감소하였다 (Fig. 6). Nq 4-6 처리구의 경우 실험 시작 시 4.1 x 105 cells mL-1 에서 7일째 3.4 x 106 cells mL-1 까지 증가하였고, 이후 실험 종료 시인 15일째 1.9 x 106 cells mL-1 까지 지속적으로 감소하였다. HNFs의 경우 대조구에서 실험 시작 시 2.5 x 103 cells mL-1 에서 10일째 1.8 x 104 cells mL-1까지 지속적으로 증가하였으며, 실험 종료 시인 15일째 1.3 x 104 cells mL-1 로 감소하였다. Nq 4-6 처리구의 경우 실험 시작 시 1.9 x 103 cells mL-1 에서 10일째 2.6 x 104 cells mL-1까지 지속적으로 증가하였으며, 실험 종료 시인 15일째 1.5 x 104 cells mL-1 로 감소하였다. Ciliates는 대조구에서 실험 시작 시 6.2 x 103 cells mL-1 에서 7일째 9.0 x 103 cells mL-1까지 증가하였으며, 이후 실험 종료 시인 15일째에 7.4 x 103 cells mL-1까지 지속적으로 감소하였다. Nq 4-6 처리구의 경우 실험 시작 시 6.2 x 103 cells mL-1 에서 4일째 8.4 x 103 cells mL-1 로 증가하였으며, 이후 실험 종료 시인 15일째에 5.6 x 103 cells mL-1까지 지속적으로 감소하였다. 식물플랑크톤을 제외한 다른 생물학적 요인은 전체적으로 유사한 경향성을 나타난 것으로 보아 Nq 4-6 화합물에 의한 영향은 미미 할 것으로 판단된다.
|
Fig. 5. Composition (%) of groups of centric diatom, pennate diatom and chlorophyceae in control (a) and treatment (b) microcosms during the experiment. |
|
Fig. 6. Changes of the abundances of bacteria, heterotrophic nanoflagellates (HNFs) and ciliates in control and treatment of naphthoquinone 4-6. |
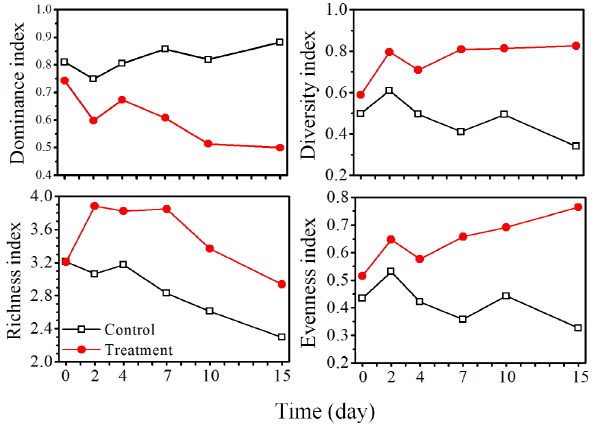
3.3 Nq 4-6에 의한 식물플랑크톤 종 다양성 변화
Nq 4-6 화합물의 미소생태 실험에서 식물플랑크톤의 다양성 지수는 대조구에서 실험 시작부터 0.80 이상으로 높았으나, Nq 4-6 처리구에서는 실험 시작 시부터 15일째 실험 종료 시까지 지속적으로 감소하였다 (Fig. 7). 우점도 지수는 처리구가 대조구보다 낮게 나타났으며, 이는 높은 현존량으로 우점한 규조 S. hantzschii가 Nq 4-6 화합물에 의한 사멸로 현존량이 감소하면서 낮아진 것으로 판단된다. 다양도 지수의 경우 대조구에서 실험 시작 시부터 15일째 실험 종료 시까지 지속적으로 감소하였고, 처리구의 경우 실험 시작 시부터 실험 종료 시까지 지속적으로 증가하였다. 전체적으로 다양도 지수는 처리구가 대조구 보다 높게 나타났다. 풍부도 지수의 경우 대조구에서 실험 시작 시부터 15일째 실험 종료 시까지 지속적으로 감소하였으며, 처리구의 경우 실험 시작 시부터 7일째까지 증가하였으며, 이후 15일째 실험 종료 시까지 감소하였다. 전체적으로 풍부도 지수는 처리구가 대조구 보다 높게 나타났다. 다양도 지수와 풍부도 지수는 대조구에 비해 처리구에서 높게 나타났다. 대조구의 경우 전체적으로 감소하는 경향을 나타냈는데, 실험초기에 대조구에서 높은 현존량으로 우점한 규조 S. hantzschii가 지속적으로 높은 현존량을 유지함으로써 다른 식물플랑크톤이 종간 경쟁에서 이기지 못해 다양도와 풍부도가 비교적 낮게 나타난 것으로 판단된다 (Morin 2011). 또한 대조구에서 인산염 인, 규산염 규소 등의 영양염 고갈로 인해 식물플랑크톤 현존량 및 종 수가 감소하면서 다양도와 풍부도가 감소한 것으로 판단된다. 처리구에서는 풍부도가 7일 이후 감소하였는데, 이는 Nq 4-6 화합물에 의해 우점종인 S. hantzschii의 현존량이 감소하고, 일부 규조류가 사멸하면서 감소하였을 것으로 판단된다. 이와 같은 결과는 앞에서 수행한 살조범위 실험 결과와 일치하는 결과로 Nq 4-6 화합물이 규조 S. hantzschii 외에 일부 규조류에 영향을 미치는 것으로 사료된다. 균등도 지수의 경우 대조구에서 실험 시작 시부터 15일째 실험 종료 시까지 증가 감소를 반복하였으며, 처리구의 경우 실험 시작 시부터 15일째 실험 종료 시까지 지속적으로 증가하였다. 균등도 지수 역시 대조구에 비해 처리구에서 높게 나타났는데, 이는 처리구에서 Nq 4-6 화합물에 의해 높은 현존량으로 우점한 규조 S. hantzschii가 감소하면서 다른 식물플랑크톤 현존량과 유사한 현존량을 나타내 균등도가 대조구에 비해 높게 나타난 것으로 판단된다.
|
Fig. 7. Dominance index, diversity index, richness index and evenness index of phytoplankton in control and treatment of naphthoquinone 4-6. |
이상의 미소생태 실험의 군집분석 결과를 종합해 볼 때, Nq 4-6 처리구에서 식물플랑크톤의 종 다양성을 증진시키는 결과를 나타냈다. 이는 Nq 4-6 화합물을 S. hantzschii가 대발생을 일으킨 수체에 적용할 경우 단순히 조류 대발생을 제어할 뿐만 아니라 수생태계 건강성에 긍정적인 영향을 미칠 것이라고 판단된다.
4. 결 론
겨울철 조류 대발생을 일으키는 규조 Stephanodiscus hantzschii를 제어하기 위해 개발된 생물유래 합성물질인 Nq 4-6 화합물을 남조 5종, 녹조 2종, 규조 4종에 적용한 결과, 규조 S. hantzschii를 포함한 원반형 규조류에 특이적으로 살조 활성을 나타냈다. 또한 현장 적용 가능성을 평가하기 위해 현장수를 이용한 미소생태 실험을 수행한 결과, Nq 4-6 화합물 적용 후 식물플랑크톤 종 다양성 및 수생태 건강성을 증진시키는 결과를 나타냈다. 결국 Nq 4-6 화합물은 S. hantzschii만을 특이적으로 제어하고 수생태 건강성을 증진시키는 친환경적 녹조 제어제로 판단된다. 하지만 본 연구는 실험실 내 실험으로 개발된 Nq 4-6 화합물을 현장 수계에 적용하기 위해서는 pilot test, test bed 등의 큰 규모에 적용해 Nq 4-6 화합물 적용 시 발생할 미소생태 변화에 대한 추가적인 연구가 필요하다.
