

1. 서 론
1.1 카메라 트랩의 발전과 활용
1.2 최소 개체수 추정 및 예측 모델링 기법의 발전
1.3 국내 연구상황과 한계
1.4 알고리즘적 접근의 가능성
1.5 본 연구의 필요성과 차별성
2. 개체수 추정 알고리즘
2.1 알고리즘 개요
2.2 이동 비용 표면 및 카메라 위치간 이동시간 산출
2.3 탐지 이벤트의 시공간적 독립성 분류
2.4그래프 분석 기반 MPS
2.5 최종 MPS 추정
3. 사례연구
3.1 대상종
3.2 연구지역
3.3 카메라 트랩 이벤트 데이터
3.4 산양 배설물의 DNA 분석으로 설정한 모집단
3.5 알고리즘 구현
3.6 최소 개채수 추정 및 시간적 변동성
3.7 결과 검증
4. 고 찰
5. 결 론
1. 서 론
1.1 카메라 트랩의 발전과 활용
카메라 트랩(camera trap)은 야생동물 연구를 위한 비침습적 모니터링 도구로서 지난 수십 년간 발전하여 다양한 목적으로 활용되고 있다(Gilbert et al. 2021). 특히 1990년대 이후 디지털 센서카메라의 도입과 기술 향상으로 카메라 트랩 연구가 급속히 증가하였으며, 1994년부터 2020년까지 관련 학술 논문의 수가 80배 이상 증가할 정도로 보편화되었다(Delisle et al. 2021). 이러한 카메라 트랩 기법의 확산은 야생동물의 서식 확인, 행동 특성 파악 등 생물다양성 모니터링의 표준 도구로 자리 잡았다.
1.2 최소 개체수 추정 및 예측 모델링 기법의 발전
개체수 추정 연구는 크게 개체 식별1)가능 종과 개체식별 불가능(이하 무표식) 종을 대상으로 하는 방법으로 구분된다. 전자는 SECR (Spatially Explicit Capture–Recapture; Efford 2004, Borchers and Efford 2008) 모델이 대표적이며, 이는 개체 탐지 위치의 공간정보를 확률 모형에 통합하여 밀도를 추정한다. 무표식 종의 경우 REM (Random Encounter Model; Rowcliffe and Carbone 2008)이나 TTE (Time-to-Event; Nakashima et al. 2018) 모델이 활용된다. REM은 무작위 이동하는 개체의 탐지율을 이용해 밀도를 추정하며, TTE는 카메라 설치 후 첫 탐지까지 걸린 시간 분포를 분석해 개체수를 산출한다.
그러나 이들 기법은 이동 패턴의 무작위성 가정, 탐지 확률 추정 불확실성, 서식지·지형 특성 반영 한계 등의 제약이 있다(Sollmann et al. 2013).
예측·모델링 측면에서, 서식지 적합도 모델(Habitat Suitability Models)과 최소비용경로(Least-Cost Path, LCP) 분석은 주로 서식지 연결성 분석과 종 이동 가능성 평가에 활용된다(Adriaensen et al. 2003; Zeller et al. 2012). 예를 들어, Dutta et al. (2016)은 서식지 적합도 모델을 바탕으로 LCP 분석을 수행하여 호랑이의 잠재적 이동 경로를 도출했고, Cushman et al. (2006)은 LCP를 사용해 곰의 이동 연결성을 평가하였다. 또한 Abrahms et al. (2016)은 LCP 결과를 이용하여 카메라 트랩 설치 지점을 최적화함으로써 목표 종의 탐지 확률을 향상시켰다. 그러나 이들 연구는 주로 카메라 배치 전략 수립이나 이동 경로 예측에 국한되며, 카메라 트랩 데이터의 시공간적 이벤트 분석과 직접 결합해 개체수를 추정한 사례는 보고되지 않았다.
1.3 국내 연구상황과 한계
국내 상황을 살펴보면, 주로 국립공원을 중심으로 야생동물 목록 조사나 서식실태 모니터링에 활용되어 왔으나, 개체군 추정과 같은 심화 연구는 상대적으로 활발하지 못하다(Park et al. 2018a, 2024, Lim et al. 2023). 한국에 서식하는 산양, 고라니, 멧돼지, 노루 등 중대형 포유류는 개체별 외형 차이가 뚜렷하지 않아 사진만으로 개체 식별 매우 어렵다. 예를 들어 산양의 경우 뿔 모양과 얼굴무늬 등 미세한 형태적 차이를 토대로 개체분류를 시도한 연구가 있었지만, 이러한 방법은 주관적이고 한계가 있다(Kim et al. 2020). 따라서 호랑이나 표범처럼 고유한 무늬로 개체를 구분할 수 없는 한국의 야생동물은 포획-재포획 방법을 적용하기 어렵고, 개체군 크기 추정을 위한 대안적 기법이 필요한 실정이다.
1.4 알고리즘적 접근의 가능성
앞서 기술한 기존 개체수 추정 기법(SECR, REM, TTE 등)은 국제적으로 널리 활용되고 있으나, 서식지·지형 특성이 개체 이동에 미치는 영향을 정량적으로 반영하지 못하거나, 개체 식별이 불가능한 종에는 적용이 제한적이라는 한계가 있다(Sollmann et al. 2013). 또한, 이동 패턴의 무작위성 가정은 종의 실제 행동 특성과 괴리가 있어, 한국과 같이 복잡한 지형과 인위적 장벽이 많은 환경에서는 정확성이 저하될 수 있다.
한국의 카메라 트랩 연구 역시 서식지 분포 조사와 활동 패턴 분석에 집중되어 있으며 개체 식별 불가능 종을 대상으로 한 정량적 개체수 추정 연구는 거의 없다. 특히, 산양과 같이 서식지가 단절되고 이동 경로가 제한적인 종의 경우, 기존 기법은 지형적 제약을 고려하지 않기 때문에 과대 또는 과소 추정 위험이 크다.
이러한 한계를 극복하기 위해서는, 서식지 이론과 공간 네트워크 분석을 결합한 새로운 접근이 필요하다.
먼저, 서식지 적합도 모델을 통해 환경변수 기반의 서식 적합도를 추정하고, 이를 이동저항표면(resistance surface)으로 변환하여 최소비용경로를 계산함으로써, 현실적인 개체 이동 가능성을 평가할 수 있다(Adriaensen et al. 2003, Zeller et al. 2012). 또한, 카메라 트랩 탐지 이벤트를 정점(vertex)으로 하고, 거리–시간 제약 조건을 만족하는 이벤트 쌍을 간선(edge)으로 연결하는 그래프를 구성하여, 연결성분 분석을 통해 최소 개체수를 추정할 수 있다(Farine and Whitehead 2015, Jacoby and Freeman 2016).
1.5 본 연구의 필요성과 차별성
이러한 문제를 해결하기 위해 본 연구에서는 카메라 트랩 이벤트 데이터2)와 서식지 기반 이동제한 요인을 결합하여 목표종의 최소 개체수를 추정하는 새로운 알고리즘을 제안하였다. 이 알고리즘의 접근 방법은 서로 다른 카메라 지점 야생동물의 최정 이동경로와 이동속도 한계를 활용하여, 임계시간 이내에 동일 개체의 이동이 불가능한 이벤트의 조합은 서로 다른 개체임을 전제로 한다.
알고리즘의 실증 적용은 설악산 오색지역의 산양을 대상으로 한 사례연구를 수행하였으며, 알고리즘의 정확성은 동일 지역의 산양 분변 DNA 분석으로 추정된 개체수를 모집단으로 설정하고, 통계적 유의성을 검증하였다.
2. 개체수 추정 알고리즘
2.1 알고리즘 개요
본 연구에서 제안하는 최소 개체수(Minimum Population Size, 이하 MPS) 추정 알고리즘은 카메라 트랩으로 수집된 시계열 데이터를 기반으로, 개체의 공간적 이동에 대한 제약 조건을 적용하여 개체수를 추정한다(Fig. 1). 알고리즘의 핵심은 두 관측 이벤트 사이의 시간 간격이 동물의 물리적 이동에 필요한 최소 시간보다 짧다면, 두 이벤트는 서로 다른 개체로 부터 발생했다는 가정을 전제로 한다. 또한, 단순한 관측빈도나 최대 동시 관측 개체수 뿐 만 아니라 동물의 이동행태와 서식지 특성을 통합한 동적 시공간 필터링(dynamic spatiotemporal filtering)기법을 적용하였다(Rowcliffe and Carbone 2008).
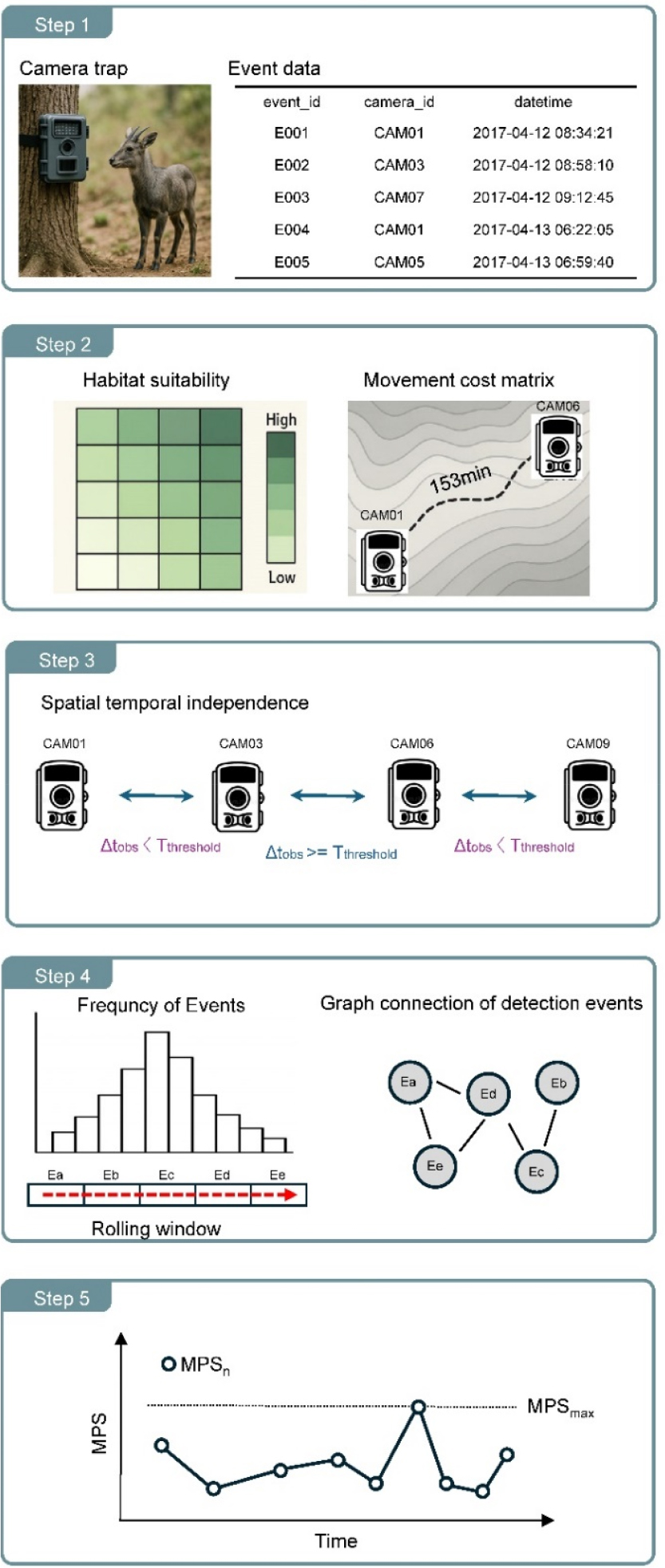
Fig. 1.
Workflow of the proposed minimum population size (MPS) estimation algorithm using camera trap event data. The process consists of five steps: (1) collecting spatiotemporal detection events from camera traps, (2) generating a movement cost matrix based on habitat suitability, (3) classifying detection events as independent or dependent based on spatiotemporal thresholds, (4) applying a rolling time window to construct graphs of connected detection events, and (5) estimating the MPS for each window and determining the final MPS as the maximum observed value.
2.2 이동 비용 표면 및 카메라 위치간 이동시간 산출
동물의 이동은 균일한 공간에서 무작위로 일어나지 않으며, 서식지 질(habitat quality)의 영향을 받는다. 서식지 질은 서식 적합도(habitat suitability)로 해석할 수 있는데, 서식 적합도가 높은 지역은 동물의 이동에 낮은 저항을 제공하는 반면, 부적합한 지역은 높은 저항을 나타낸다. 따라서 서식지에 대한 선호도는 야생동물의 이동 경로와 속도를 결정하는 핵심 요인이 된다(Spear et al. 2010).
본 알고리즘에서는 종분포모델(species distribution models)을 통해 서식지 적합도를 공간 데이터인 래스터(raster) 형태로 산출한다. 공간 좌표 (x,y)에서의 서식지 적합도를 H(x, y)라고 정의할 때, 이동 비용 C(x,y)는 다음과 같이 H(x, y)의 역수로 계산된다. 이는 동물이 선호하는 서식지일수록 서식지 적합도가 높아지고, 그에 따라 이동 저항은 낮아져 결과적으로 이동비용이 감소함을 의미한다(Eq. 1).
여기서, ε는 서식 적합도가 0일때 분모가 0이 되는 것을 방지하기 위한 상수
이동 비용 표면 위에서, 두 카메라의 위치 i와 j를 연결하는 가능한 경로 중 누적 비용이 가장 적은 최소 비용 경로(Least cost path)를 산출하며, 이 경로의 총비용은 Dcost(i,j)로 정의한다.
산출된 비용(Dcost)은 동물의 평균 이동 속도()를 이용해 두 지점 간 최소 이동 시간(Minimum Travel Time) T(i,j)로 변환한다(이후 과정의 수식 간결성을 위해 T(i,j)를 𝜏로(Eq. 2) 정의한다).
2.3 탐지 이벤트의 시공간적 독립성 분류
이 과정은 알고리즘의 핵심으로, 시공간적으로 인접한 두 탐지 이벤트(Ea, Eb)가 동일 개체로 부터 발생할 가능성을 평가한다(O’Brien et al. 2003). 만약 한 개체가 물리적으로 이동하는데 소요되는 시간보다 짧은 시간 안에 두 지점에서 관측되었다면, 이는 서로 다른 개체로 판단한다.
여기서 비교시간은 임계시간(Tthreshold)으로 정의하고, 이벤트가 발생한 카메라 ID가 동일한지 여부에 따라 다르게 설정된다(Eq. 3).
여기서, 𝜏buffer는 동일 카메라 내에서 독립적인 이벤트로 간주하기 위한 최소 기간 간격
𝜏i,j는 카메라 i와 j사이의 최소 이동시간
위의 관측 시간의 차이(∆tobs)와 앞서 정의한 를 비교하여 이벤트를 종속 이벤트(Dependent Events)와 독립 이벤트(Independent Events)로 분류한다.
종속 이벤트는 만약 ∆tobs ≥𝜏이면, 한 개체가 i에서 j로 이동하여 두 이벤트를 생성했을 가능성이 충분하다. 따라서 두 이벤트는 동일 개체에서 비롯된 종속 관계로 분류한다. 반면, 독립 이벤트는 ∆tobs <𝜏의 조건을 만족하며, 한 개체가 물리적으로 임계 시간안에 두 지점을 이동하는 불가능하므로, 두 이벤트는 서로 다른 개체에 의한 독립관계로 분류한다(Eq. 4).
여기서, IC는 Independence classification
E는 탐지 이벤트
2.4그래프 분석 기반 MPS
시공간적에서 종속적3)으로 연결된 탐지 이벤트의 집합은 한 개체의 활동으로 간주할 수 있다(Whitehead 2008). 따라서, 독립성 분류 결과(Eq. 4)를 바탕으로, 이벤트들을 연결하여 개체 별 시공간적 활동 궤적4)을 재구성하고, 이를 통해 MPS를 산출한다(Jacoby and Freeman 2016).
여기서 전체 연구기간을 k일 단위로 개념의 시간 창(time window)5)를 적용하여 롤링 윈도우(rolling window)6) W로 분할한다(Holme and Saramäki 2012).
각 창(W) 내에서, 모든 탐지 이벤트를 정점(vertex)으로 설정하고, (Eq. 4)에 따라 종속 관계인 두 이벤트를 간선(edge)으로 연결하여 무방향성 그래프7) 를 구축할 수 있다.
GW 그래프에서 서로 연결된 정점들의 집합인 연결요소(connected component)의 총개수를 해당 시간 창의 최소 개체수로 정의할 수 있으며(Farine and Whitehead 2015), 각 연결 요소는 한 개체의 시공간적 활동 궤적으로 해석할 수 있다(Eq. 5).
여기서, MPSW는 특정 시간 창(W)내의 최소 개체수
GW는 해당 시간 창에서 종속 탐지 이벤트를 네트워크로 연결한 그래프
C(GW)는 그래프(GW)가 완전히 분리된 연결 요소의 개수
2.5 최종 MPS 추정
야생동물 개체군의 활동성은 계절, 시간, 사회적 상호작용 등 다양한 요인에 따라 유동적이므로, 특정 개체가 카메라에 탐지될 확률은 일정하지 않다(Rowcliffe et al. 2014). 따라서 단일 시간 창에서 산출된 MPS 값은 해당 기간의 탐지 확률에 크게 의존하며, 전체 개체군을 대표하지 못할 수 있다(Sollmann et al. 2013).
그러므로, 각 시간 창에서 계산된 MPSW는 해당 기간에 존재가 확실하게 증명된 개체들의 최솟값이다. 실제 개체수 N은 항상 N≥MPSW관계를 만족하기 때문에 MPSW는 실제 개체수의 하한선을 나타내는 보수적인 추정치이다(Eq. 6).
결론적으로 최종 MPS는 개별 시간 시창에서 산출된 모든 최소 개체수의 최대값으로 결정된다. 이 로직은 카메라 트랩 탐지 기간 중 탐지율이 가장 높아 가장 많은 수의 개체가 동시에 식별된 시점의 정보를 반영한 것이다.
3. 사례연구
3.1 대상종
연구 대상종은 산양(Long-tailed goral, Naemorhedus caudatus)이다(Fig. 2). 산양은 한국, 중국 동북부, 러시아 극동 일부 지역에 분포하는 산악성 반추동물로, 주로 해발 500–1,500 m의 급경사 암벽 지대에 서식한다. 암석과 절벽이 혼재하는 지형은 포식자로부터의 회피와 번식지 확보에 유리하며, 주요 먹이는 관목, 풀, 나뭇가지, 이끼류로 계절에 따라 식이 구성이 변한다. 산양은 주로 새벽과 황혼 무렵에 활동하는 주행성(crepuscular) 경향을 보이며, 영역성이 뚜렷하고 연중 서식지를 크게 이동하지 않는 정주성을 나타낸다(Lee et al. 2019).

한국에서는 설악산국립공원을 비롯한 백두대간 산악 지대에 분포하며, 대부분 절벽과 능선부를 연결하는 좁은 이동 경로를 이용한다. 이러한 생태적 특성은 서식지 단절과 인위적 교란에 매우 민감하게 작용하며, 특히 도로, 스키장, 채석장 등 인프라 개발은 산양 개체군의 연결성을 저해하는 주요 요인이다(Lee et al. 2019).
본 연구의 사례 지역인 설악산국립공원은 산양의 주요 핵심 서식지 중 하나로, 급경사 암벽과 혼효림이 발달하여 연중 산양의 서식에 적합한 환경을 제공한다.
산양은 계절 털갈이와 암수 성적 형태 차이가 분명하지 않아 사진만으로 개체 식별이 어렵다. 또한 산양은 급경사 암벽과 협곡을 이용하는 특수한 서식 특성을 가지고 있기 때문에 카메라 트랩 설치 공간이 지형적으로 제한되어 탐지 이벤트의 위치 불확실성이 적고(Park et al. 2018b), 서식 적합도 기반 비용-표면을 구축할 때 베타 다양성의 해석이 명확하다는 장점이 있다.
3.2 연구지역
연구지역은 설악산국립공원 오색권역(38°45′–38°57′ N, 128°25′–128°28′ E)으로, 설악산의 최고봉인 대청봉(해발 1,708 m)의 남서 사면이다. 연구 경계는 고도 270~1,705 m 범위에 걸쳐 있으며, 총 면적은 7.7 km2으로 77개의 카메라 트랩이 배치되었다(Fig. 3).
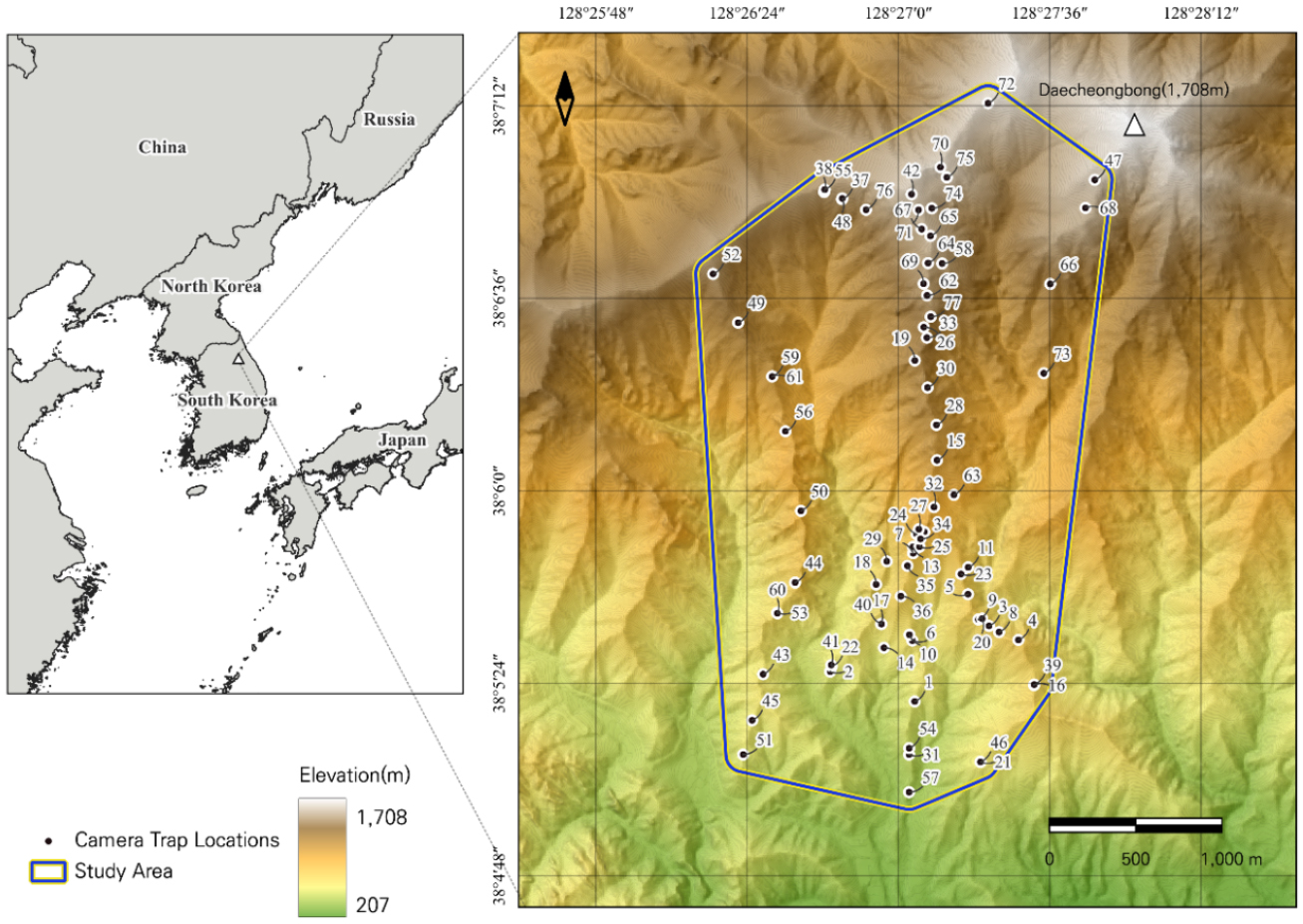
Fig. 3.
This figure shows the study area and camera trap locations. The case study area is Seoraksan National Park, specifically the southwestern slope of Daecheongbong (1,708 m a.s.l.), the highest peak of Seoraksan Mountain. The study boundary spans an elevation range of 270–1,705 m, covering a total area of 7.7 km2. A total of 77 camera traps were deployed in this area at approximately 1 km intervals.
3.3 카메라 트랩 이벤트 데이터
카메라 트랩의 운영 기간은 2016년 10월 27일부터 2017년 12월 21일로 총 420일이며, 산양의 이벤트가 탐지된 카메라는 총 46대이다. 이벤트 탐지 건수는 1,185건이로 카메라 1대당 평균 25.8건을 탐지하였고, 중앙값은 11건이다. 최다 탐지 카메라는 CAM59 (192건), CAM57 (187건) 이고, 최소 탐지는 CAM36, 04, 24 등으로 각 1건이다(Fig. 4).
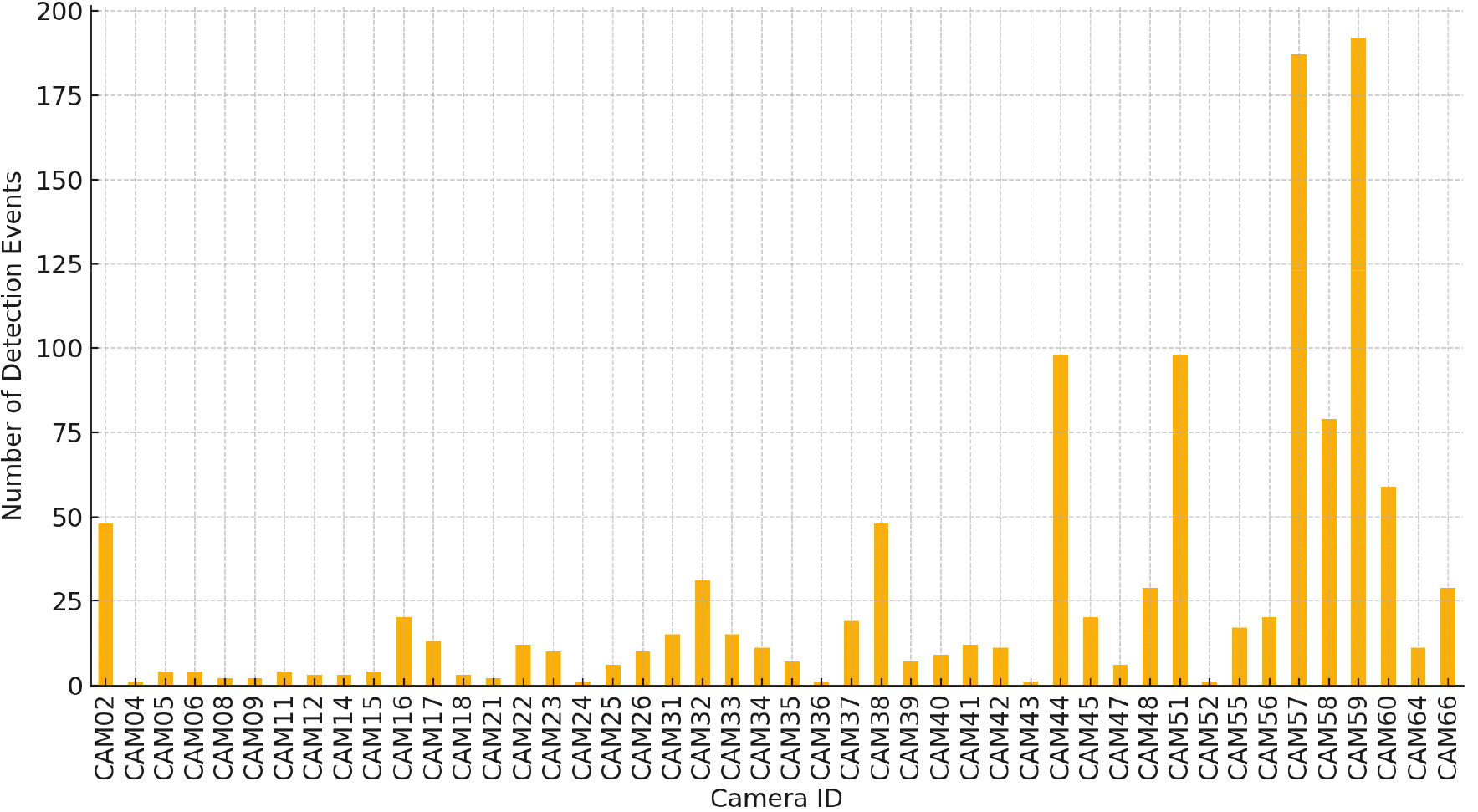
Fig. 4.
Camera traps were operated for 420 days. Although 77 cameras were deployed, long-tailed goral events were detected by 46 cameras. A total of 1,185 detection events were recorded, with an average of 25.8 detections per camera and a median of 11 detections. The cameras with the highest detection rates were CAM59 and CAM57, while those with the lowest detection rates included CAM36, CAM04, and CAM24.
3.4 산양 배설물의 DNA 분석으로 설정한 모집단
본 연구의 알고리즘 검증을 위해 산양 개체군의 모집단은 25개체로 가정하였다. 이 값은 Kim et al. (2025)의 연구 결과로, 2019년, 2022년, 2023년에 분변 112점을 mtDNA control region 서열을 활용해 개체를 식별하고, 오색권역(남서사면)에서 25개체(95 % CI = 21–29)를 추정한 것이다.
본 연구의 카메라 트랩 운영기간과 개체수 연구의 DNA 채집기간은 약 5년의 차이가 있다. 그러나 설악산 산양의 연간 순증식률은 6% 이내로 제한되어 있어, 5년 이내의 개체수 변동은 한 자리 수 범위에 머물 것으로 예상된다(Jang et al. 2020). 또한 GPS collar 추적 결과에 따르면, 산양은 좁은 행동권(≤1 km2)과 짧은 일일 이동거리(≈1 km)를 갖는 정주적(sedentary) 특성을 보이며, 서식지 선택이 암벽·급경사 지형에 집중되는 것으로 나타났다(Cho et al. 2014).
이러한 생태적 특성을 고려할 때, 카메라 트랩 운영과 분변 채집기간의 시간적 차이가 알고리즘을 적용한 최소 개체수 비교에 유의미한 편향을 발생시키지 않을 것으로 판단하여 알고리즘에서 추정한 개체수의 통계적 검증을 위한 모집단으로 사용하였다.
3.5 알고리즘 구현
본 사례연구의 MPS 추정 알고리즘은 RStudio (IDE 2023.09.1+494)에서 R 4.3.2 환경으로 수행하였다. 알고리즘 구현의 흐름은 (i) 공간 비용-표면 구축, (ii) 최소 이동시간 행렬 산출, (iii) 탐지 이벤트 독립성 판정, (iv) 창(window)별 그래프 분석, (v) 최소 개체수(MPS) 산정으로 구성되며, 사용 패키지와 함수는 Table 1과 같다.
Table 1.
R packages used for each analytical step
| Step | Main packages (version) | Key functions / objects | Reference |
| Cost-surface generation | raster 3.6-23 | raster(), calc() | (Hijmans 2018) |
| Transition matrix & resistance distance | gdistance 1.6-4 | transition(), geoCorrection(), costDistance() | (Van Etten 2017) |
| Spatial‐vector processing | sf 1.0-14 | st_as_sf(), st_transform() | (Pebesma 2018) |
| Data wrangling | dplyr 1.1-3; tidyr 1.3-0; lubridate 1.9-3; readr 2.1-4 | mutate(), crossing(), ymd_hms() | (Rathod et al. 2023) |
| Graph analysis | igraph 1.5-1 | graph_from_data_frame(), components() | (Csardi & Nepusz 2006) |
| Visualisation | ggplot2 3.4-4 | geom_col(), geom_line() | (Villanueva and Chen 2019) |
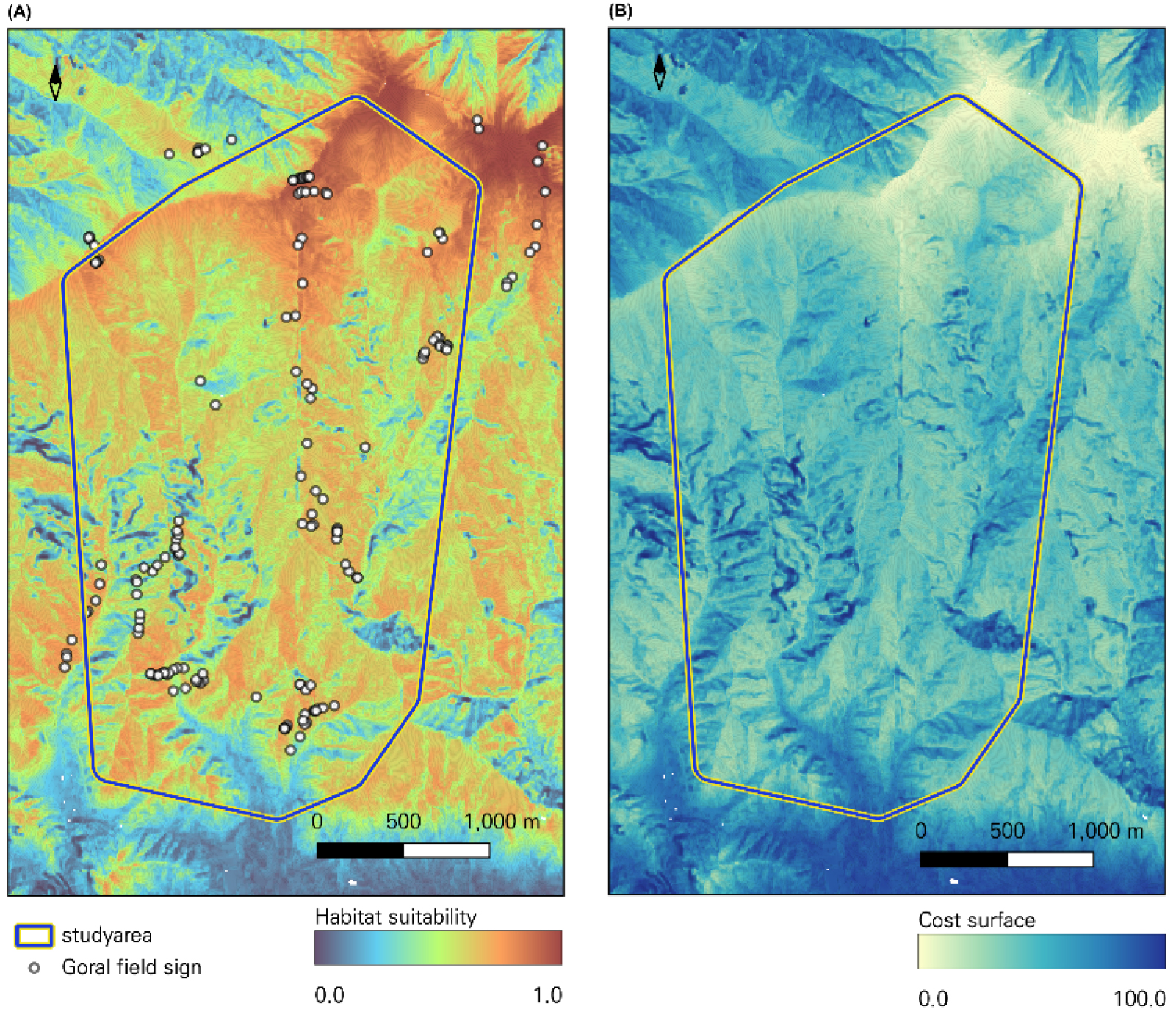
Fig. 5.
(A) shows the habitat suitability results for goral using species distribution modeling. White dots represent goral habitat signs used as dependent variables in the species distribution model, consisting primarily of fecal pellets. (B) presents the conversion of the results from (A) into a cost surface using Eq (1), which served as the baseline data for analyzing movement resistance between camera trap locations.
3.5.1 공간 비용-표면 및 이동시간 행렬
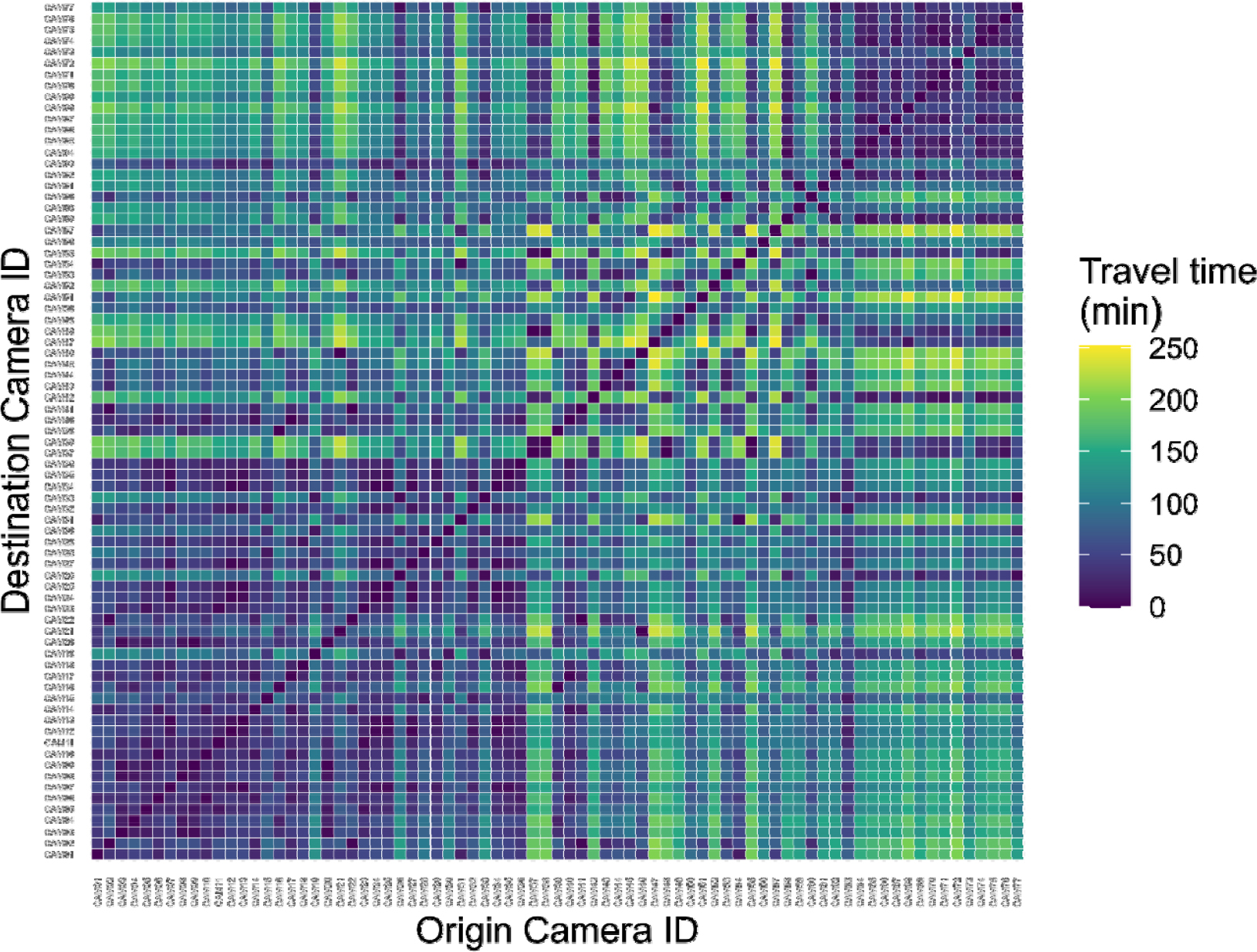
공간 비용-표면은 종분포모델을 활용하여 산양의 서식 적합도를 산출하고 (Eq. 1)을 적용하여 비용표면으로 전환하였다(Fig. 5). 이후 카메라 정점 조합별 저항거리를 계산하고, 산양의 평균 보행속도 로 적용하여 이동시간 행렬을 도출하였다(Fig. 6).
𝜐는 기존 산양 추적 연구에서 보고된 일일 이동 거리(5–10 km/day)를 기반으로 활동 시간대에 대한 보정을 통해 추정한 값이며(Cho et al. 2014, 2015), REM (Random Encounter Model) 및 시간기반 알고리즘 적용 시의 일반적인 이동속도 가정 범위와 부합한다(Gray 2018).
3.5.2 탐지 이벤트의 종속과 독립성 분류
연구 기간 동안 수집된 모든 탐지 이벤트 간의 모든 쌍을 구성하고, 각 쌍에 대해 관측된 시간 차이(∆t)와 서식지 저항을 고려한 최소 이동 시간(𝜏)을 계산하였다. 이 두 변수 간의 관계를 기반으로 각 이벤트 쌍을 동일 개체에서 발생했을 가능성을 종속(Dependent)로, 물리적으로 불가능하고 서로 다른 개체로 간주되는 독립(Independent) 그룹으로 분류하였다.
동일 카메라의 임계시간(Eq. 3)은 카메라 트랩 설정 값에 따라 달라지며 본 연구에서 𝜏buffer는 60초로 설정하였다. 그 이유는 카메라 셋팅에 의한 연속촬영을 하나의 이벤트로 통합하기 위한 최소간격으로 본 연구의 촬영 셋팅은 이벤트 발생 시 10초 간격으로 연속촬영(5매)하고, 다음 촬영까지 트리거 인터벌이 10초이기 때문이다.
분류 알고리즘의 타당성을 시각적으로 검증하기 위해, 각 그룹에서 동일한 수의 이벤트 쌍을 무작위로 추출하고 산점도로 시각화 하였다(Fig. 7). 산점도의 x축은 실제 관측된 시간차(∆t), Y축은 이론적인 최소 이동시간(𝜏)을 로그 스케일로 나타낸 것으로 적색 점선은 ∆t=𝜏가 성립하는 이론적 경계선이다.
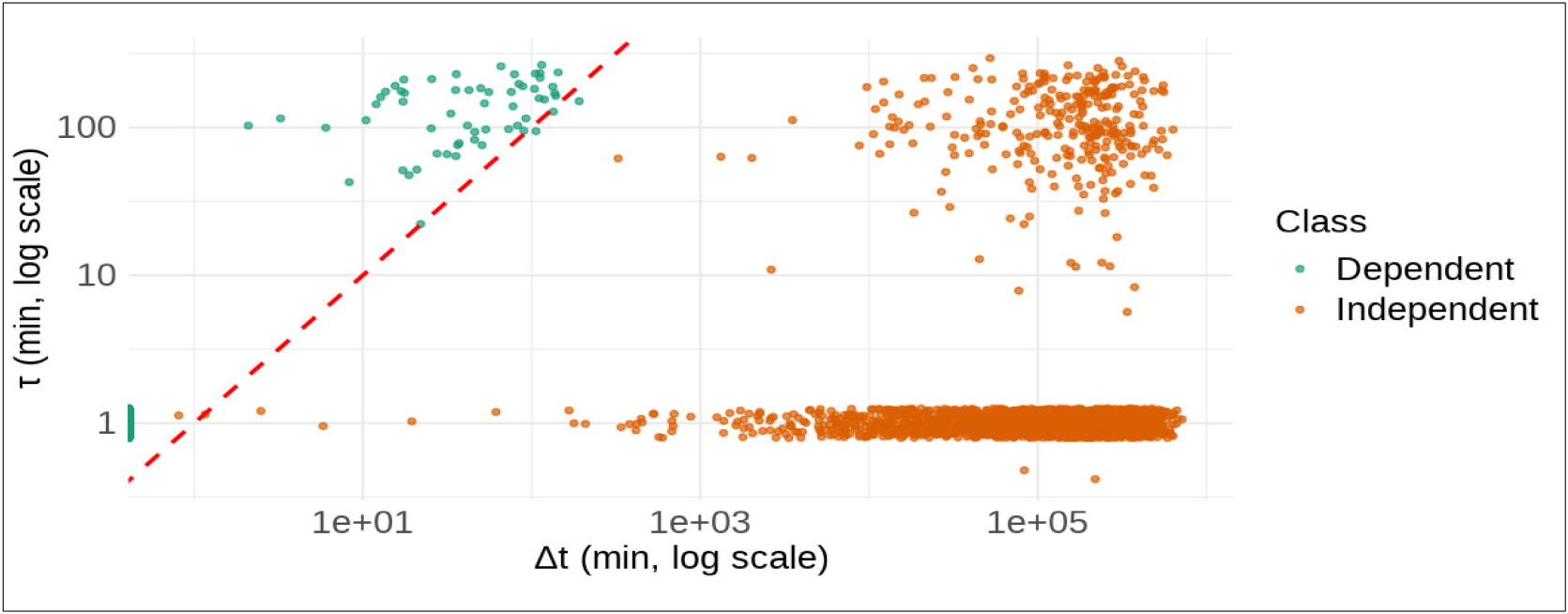
Fig. 7.
Relationship between observed time difference (Δt) and minimum travel time (τ) for event pairs. The red dashed line (Δt=τ) represents the theoretical classification boundary, clearly showing the classification results into 'dependent' (teal) and 'independent' (orange) categories based on this threshold. Data were randomly subsampled with equal numbers from each group and displayed on log-log axes for visual clarity.
분류 결과, 한 개체가 두 지점 사이를 이동하는데 필요한 최소시간(𝜏) 보다 실제 관측된 시간차 ∆t가 짧아서 물리적으로 불가능한 이벤트 쌍, 즉 𝜏>∆t조건은 모두 경계선 위쪽에 위치하며 독립으로 분류되었다. 반면, 이동에 충분한 시간이 확보된 이벤트 쌍 𝜏≤∆t 조건의 그룹은 모두 경계선 아래쪽에 위치하여 종속으로 분류되었다.
3.6 최소 개채수 추정 및 시간적 변동성
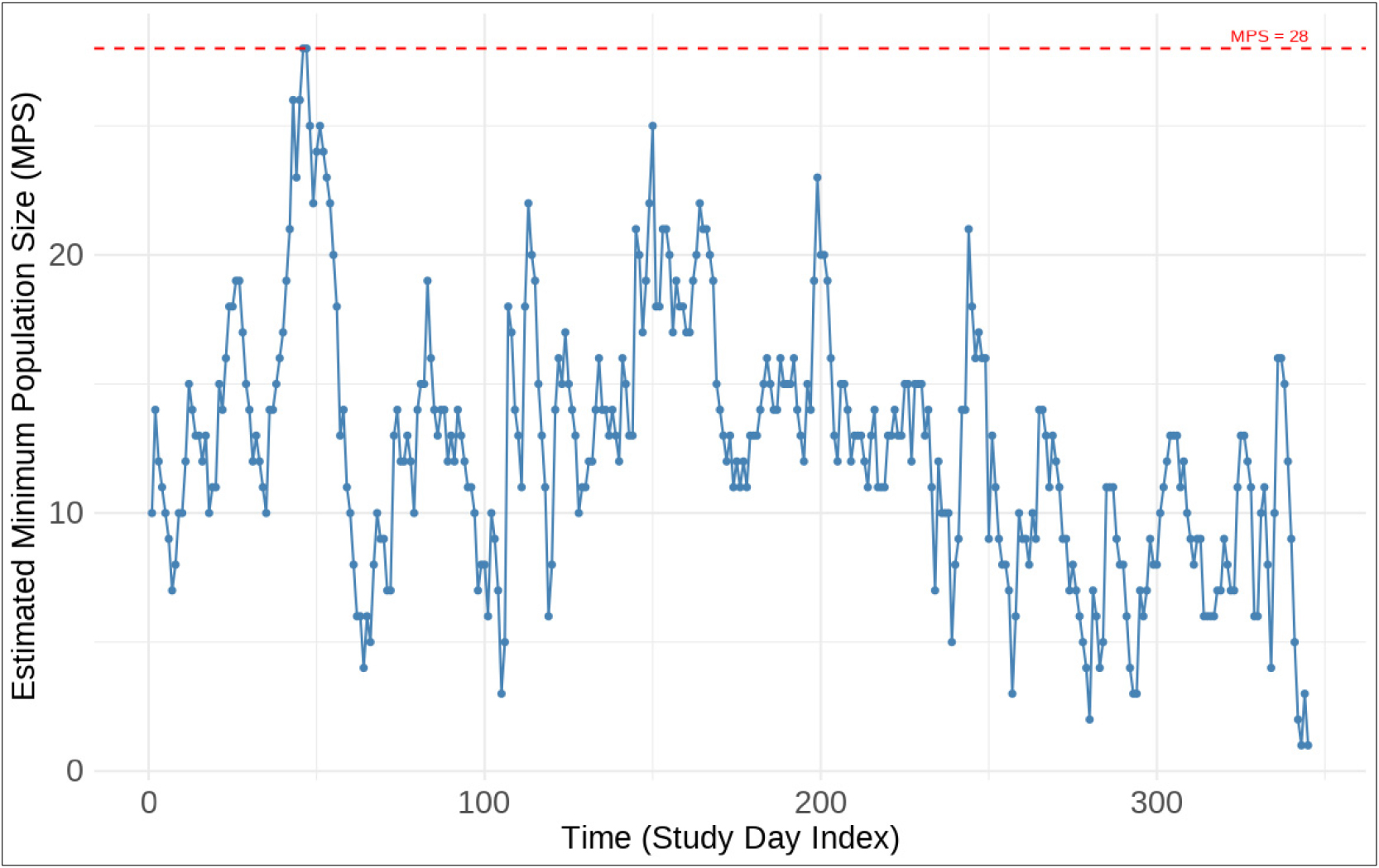
Fig. 8은 연구 기간 동안 창(rolling window) 방식으로 추정된 MPS의 시간적 변화를 나타낸 것이다. MPS는 시계열에 따라 변동폭이 크며, 최저 1개체에서 최고 28개체로 나타났다. 따라서, 본 연구에서 MPS 는()는 전체 기간의 MPS 값의 최대값으로 28개체이다.
3.7 결과 검증
본 연구에서 알고리즘으로 추정된 MPS가 개체군 모집단으로 정의한 25개체와 통계적으로 양립 가능한지 검증하기 위해서 푸아송 분포를 사용하였다. 푸아송 분포는 특정 시간 또는 공간 단위 내에서 발생하는 사건의 횟수(count) 모델링에 특화된 이산확률분포이다. 본 연구에서 알고리즘을 통해 추정한 MPS는 연구 지역내 존재하는 동물의 개체수를 나타내는 계수 자료이므로, 푸아송 분포의 기본 가정에 부합한다.
푸아송 검증의 귀무가설은 “연구지역의 산양 개체수는 25개체이다(H0: N=25)”로 설정하였다. 알고리즘으로 추정된 개체수 28에 대하여 28개체 이하로 관측될 누적 확률을 계산한 결과 p-value는 0.763으로 나타났다. 이 값은 일반적 유의수준(α=0.05)보다 상당히 크므로 귀무가설을 채택하였다(Fig. 9).
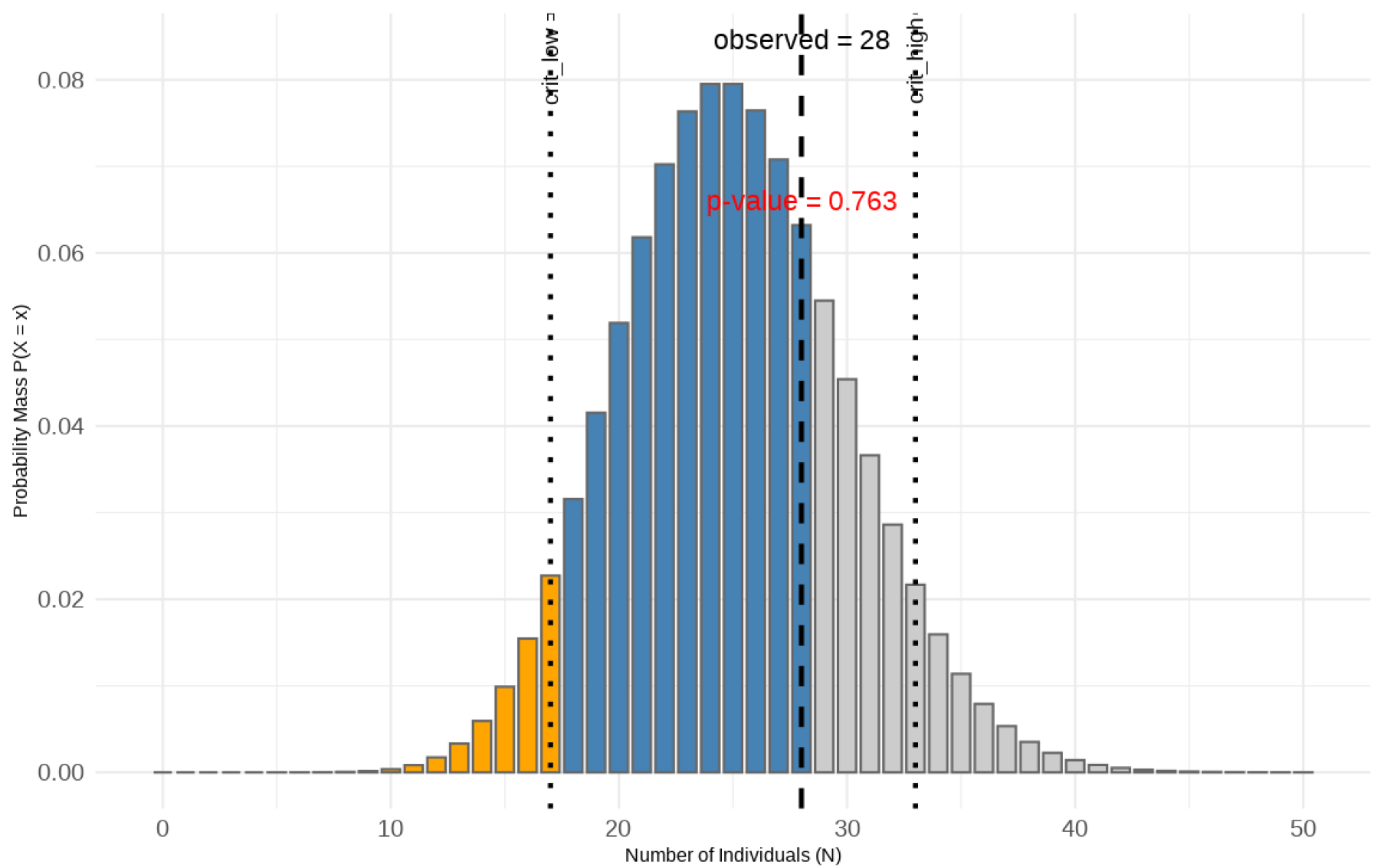
Fig. 9.
Poisson distribution comparison between observed minimum population size (MPS = 28) and null hypothesis (H₀: N = 25). The observed value lies within the confidence interval of the null hypothesis, and the p-value (0.763) exceeds the significance level (0.05), demonstrating no statistically significant difference.
결론적으로, 본 알고리즘에서 추정한 28개체는 연구지역의 개체군 모집단 25개체와 통계적으로 유의한 차이를 보이지 않는 것으로 나타났다. 이는 자연 개체군에서 충분히 일어날 수 있는 범위 내의 변동으로 해석되며, 따라서 본 연구에서 제안한 알고리즘이 실제 개체군 크기를 적절히 추정할 수 있는 유효한 방법임을 확인하였다.
4. 고 찰
본 연구는 카메라 트랩 데이터를 기반으로 개체 식별이 어려운 야생동물의 최소 개체군을 추정할 수 있는 새로운 알고리즘을 제안하였다. 본 연구는 거리-시간 제약을 이용해서 서로 동일 개체일 수 없는 이벤트를 체계적으로 구분하고, 그래프 이론을 통해 이벤트 간 관계를 구조화하여, 개체 식별이 불가능한 조건에서도 신뢰성 있는 MPS 추정을 가능하게 한다.
기존의 카메라 트랩 기반 개체군 추정 연구들은 대부분 개체 식별이 가능한 야생동물을 대상으로 한다(O’Connell et al. 2011). 호랑이의 줄무늬나 표범의 반점 무늬처럼 개체별 패턴이 명확할 경우, 전통적인 포획-재포획 기법을 적용하여 매우 높은 정확도로 밀도 및 개체군 크기 추정이 가능하다(Karanth and Nichols 1998). 그러나 우리나라에 서식하는 노루, 산양, 멧돼지, 삵 등과 같이 외형적으로 개체별 식별이 어려운 종에는 이러한 기법의 적용이 근본적으로 불가능하다.
이러한 한계를 극복하기 위해 개체 식별이 필요 없는 대표적인 방법론인 Random Encounter Model (REM)이 제안되었다. REM은 카메라의 탐지 빈도, 동물의 이동 속도 등의 정보를 이용하여 밀도를 추정할 수 있다(Rowcliffe and Carbone 2008). 하지만 이 모델은 동물이 무작위로 이동한다는 핵심 전제를 포함하며, 평균 이동 속도, 탐지 각도, 탐지 거리 등 현장에서 정확한 측정이 어려운 다양한 매개변수를 요구한다. 이러한 전제가 충족되지 않거나 매개변수 추정이 부정확할 경우, 결과의 신뢰도가 크게 저하될 수 있다는 한계가 지적되어 왔다(Iijima 2020).
기존의 포획-재포획 및 이를 응용한 공간 포획-재포획(Spatially Explicit Capture-Recapture, SECR) 모델 역시 개체 식별이 전제된다. 또한, 최근 제안된 대안적 모델인 시간 모델(Time-to-Event 또는 Waiting Time Model) 역시 불확실한 파라미터에 의존하고 있다(Santini et al. 2022, Howe et al. 2022). 이와 달리 본 연구에서 제안하는 알고리즘은 거리-시간 제약에 의한 “물리적으로 동일 개체일 수 없는 이벤트 쌍”을 판별하고 그래프 이론을 이용하여 연결성분을 구성함으로써 MPS를 추정한다. 개체 식별이 전혀 필요하지 않으면서도 기존 방법론들이 의존하는 불확실한 파라미터를 최소화했다는 점에서 기존의 개체군 추정 기법과 명확히 구분된다.
5. 결 론
본 연구에서 제안한 알고리즘의 결과를 검증하기 위해 채택한 비교 방법은 배설물 DNA 분석에 의한 개체수 추정이다. 이 방법은 개체 식별이 불가능한 종에서도 개체 고유의 유전자 마커를 이용하여 개체를 구분할 수 있으며(Waits and Paetkau 2005), 카메라 트랩 탐지 자료와 비교 가능한 수준의 공간·시간 범위에서 적용이 가능하다. 특히, 본 연구 지역의 산양 개체군은 기존 연구를 통해 배설물 DNA 기반 개체수 데이터가 축적되어 있어, 동일 시기·동일 지역의 데이터를 비교함으로써 외부 환경 변화나 서식지 변동이 결과에 미치는 영향을 최소화할 수 있었다. 따라서, 본 연구에서 채택한 배설물 DNA 분석은 자료 가용성과 방법론적 신뢰성 측면 모두에서 알고리즘 검증에 적합하다.
본 연구에서 제안한 알고리즘을 적용하기 위해서는 몇 가지 전제 조건이 필요하다. 첫째, 대상종의 이동속도에 대한 신뢰성 있는 정보가 확보되어야 한다. 이동속도는 거리-시간 제약을 적용하는 핵심 기준으로, 과대 또는 과소 추정되면 알고리즘에서 이벤트 쌍의 종속, 독립 결정 기준이 왜곡될 수 있다. 따라서 GPS 추적, 관찰 기록 등 다양한 수단을 통해 대상종의 평균 이동속도 및 최대 이동거리 범위에 대한 연구가 선행되어야 한다(Sollmann et al. 2012, Rowcliffe et al. 2014).
둘째, 서식지 기반 최소비용경로 분석을 활용하기 때문에 고품질의 종분포모형이 구축되어야 한다. 최소비용경로는 서식지 적합지수의 역수를 동물의 이동 저항값으로 활용한다(Zeller et al. 2012). 종분포모델은 종 출현데이터를 종속변수로, 지형, 식생, 기후 등의 환경요인을 독립변수로 설정하여 모델링하게 된다. 이 모델이 부정확하면 이동시간 행렬 자체의 오류를 유발하게 되어 알고리즘의 기본 전제가 흔들리게 된다(Elith and Leathwick 2009).
셋째, 본 알고리즘이 산출하는 개체군은 보수적 하한선인 “최소 개체군”이기 때문에 실제 개체군 크기와는 차이가 있을 수 있다. 예를 들어 본 연구에서 창(window) 단위 분석을 통해 산양의 최소 개체수를 28개체로 추정하였고, 이는 유전학적 추정치인 25개체와 통계적으로 유의한 차이를 보이지 않았다. 하지만 이 값은 어디까지나 최소 개체수이며, 절대 개체수 추정과는 개념적으로 구분될 필요가 있다(Otis et al. 1978).
이상의 결과를 종합하면, 본 알고리즘은 개체 식별이 어려운 종에 대해서도 최소 개체군을 신뢰성 있게 추정할 수 있는 가능성을 제시하였다. 특히 국내와 같이 야생동물의 주요 서식지인 산악지형에서 효율적인 카메라 트랩 설치만으로 개체군 정보를 취득할 수 있음을 보였다. 또한 본 알고리즘을 응용하여 행동권 분석, 공간분포 해석, 서식지 단편화 평가 등 다양한 보전연구에 확장 적용할 수 있는 토대를 제공할 것으로 기대한다.
