

1. 서 론
2. 연구 방법
2.1 조사지역 및 시료 채집
2.2 배설물시료의 DNA 추출 및 염기서열증폭
2.3 차세대염기서열분석(Next Generation Sequencing)
2.4 데이터 분석
3. 결 과
3.1 섭식 먹이원 구성
3.2 먹이원의 계절별 차이
4. 고 찰
5. 결 론
1. 서 론
고라니(Hydropotes inermis)는 포유강(Mammalia), 우제목(Artiodactyla), 사슴과(Cervidae)에 속하는 동물로 국제자연보전연맹(International Union for Conservation of Nature, IUCN)에 멸종위기 “취약(vulnerable)” 등급으로 분류되어져 있다(Harris and Duckworth 2015). 주로 중국과 한국에 분포하는 토착종이며 이 두 군집은 지리적 분포와 체색을 기준으로 두 아종(H. inermis inermis, H. inermis argyropus)으로 구분된다(Schilling and Rössner 2017). 중국 개체군은(H. inermis inermis) 서식지 파편화와 밀렵으로 인해 양쯔강 이남의 저우산 군도와 상해 인근으로 서식 반경이 축소되어 지속적인 보전 노력이 수행되고 있는 중이나(Hu et al. 2006, Li et al. 2023), 국내 개체군(H. inermis argyropus)의 경우 한반도 내 높은 서식 밀도를 보이며 경작지 등에 출현하여 농작물에 피해를 주고 있어 환경부에서는 고라니를 「야생생물 보호 및 관리에 관한 법률」에 따른 유해야생생물로 지정하여 개체수를 조절하고 있다(Park et al. 2008, Ministry of Environment 2025). 지역 별 고라니 기초생태에 대한 충분한 과학적 데이터를 바탕으로 농작물 피해가 높은 지역에 대한 방제 전략을 수립하거나 개체수 조절을 진행해야 할 필요가 있으나 국내에서는 아직 충분한 연구가 진행되지 않아 합리적인 대응책 수립에 한계가 있다.
고라니의 기초 생태를 알기 위해서는 먹이원 연구와 행동생태에 대한 연구가 핵심적이며, 특히 먹이원의 경우 고라니의 하루 활동 시간 중 섭식활동이 50.4%를 차지하여 해당 종의 주된 행동 반경과 이동을 결정짓는 중요한 요인이다(Zhang 2000). 국내에서 진행된 행동권 연구에서도 고라니가 음수를 위해 하천이나 습지와의 거리가 1.6 km 이내인 지역을 선호하는 경향이 있으며(Kim et al. 2011b), 고라니 행동권의 지역적, 계절적 차이가 먹이를 섭식하기 위한 것에 기인하는 것으로 판단하고 있다(Park and Lee 2013, Kim et al. 2021a). 국외의 선행연구에서는 고라니를 농후식섭식자(concentrate selectors, CS) 또는 중간섭식자(intermediate opportunistic feeders, IM)로 주로 관목이나 나무의 잎, 새싹 등 연한 부분을 섭식하는 browsing과 함께 과실 또한 섭식한다고 밝혔으나(Hofmann 1989), 국내 연구에서는 서식환경과 계절에 따라 주요 먹이원에 편차가 큰 것으로 나타났다. 강원도 철원과 전라남도 동부에서는 초본류(forbs, 68.85%)와 벼과류(graminoids, 30.23%)를 선호하는 것으로 나타났으며(Park et al. 2011), 산림 지역에 비해 습지 지역에서 수행된 연구에서 초본류를 더 많이 섭식하는 것으로 나타났다(Kim et al. 2011a, Kim et al. 2021b, Lee et al. 2022). 반면, 산악 지형에서 수행된 고라니 연구의 경우 활엽수림의 이용률이 높은 것으로 보고되었다(Kim et al. 2011b, Park and Lee 2013, Kim et al. 2021a, Lee et al. 2022). 따라서 고라니의 먹이 섭식은 서식지 유형에 따라 달라질 수 있으며, 서식지 및 계절적 특성을 반영한 고라니 섭식에 대한 연구가 요구된다.
최근에는 차세대염기서열분석법(next generation sequencing, NGS)을 이용한 비침습적 먹이원 연구가 활발하게 진행되고 있다(Hawlitschek et al. 2018, Ando et al. 2020, Joo et al. 2023). 차세대염기서열분석방법을 이용하여 배설물로부터 섭식된 먹이원을 검출하는 방법은 소화된 배설물 내에 짧은 절편 상태로 존재하는 DNA를 민감하게 탐지할 수 있어 metabarcoding 연구에 적합하며, 생어 시퀀싱(Sanger sequencing)과 달리 cloning 과정 없이 동시에 많은 종을 탐지할 수 있어 상대적으로 비용이 절감된다는 장점이 있다(Swift et al. 2018, Gong et al. 2019, Gueuning et al. 2019). 또한 위내용물 분석은 소화과정에서의 잔유물에 대한 형태적 분석에 기반하여 먹이원 동정의 해상도가 목 수준으로 낮고 연구자의 높은 사전지식을 요구하는 반면, 염기서열을 기반으로 한 metabarcoding 연구는 보다 높은 해상도의 먹이원 동정이 가능하며 식물분류에 대한 사전 지식 없이도 분석이 가능하다는 장점이 있다(Jang‐Liaw 2021, Liu et al. 2021). 따라서 본 연구에서는 설악산국립공원 내 국가장기생태연구지소인 점봉산 지역에 서식하는 고라니를 대상으로, 차세대염기서열분석법을 적용한 배설물 내 먹이원 분석을 통해 산림 지역 내 서식하는 고라니에 대한 먹이원 이용을 파악하고자 한다. 본 연구의 결과는 추후 고라니의 관리 대책 수립이나 보전정책 수립 시 기초 생태 데이터로써 활용될 수 있다.
2. 연구 방법
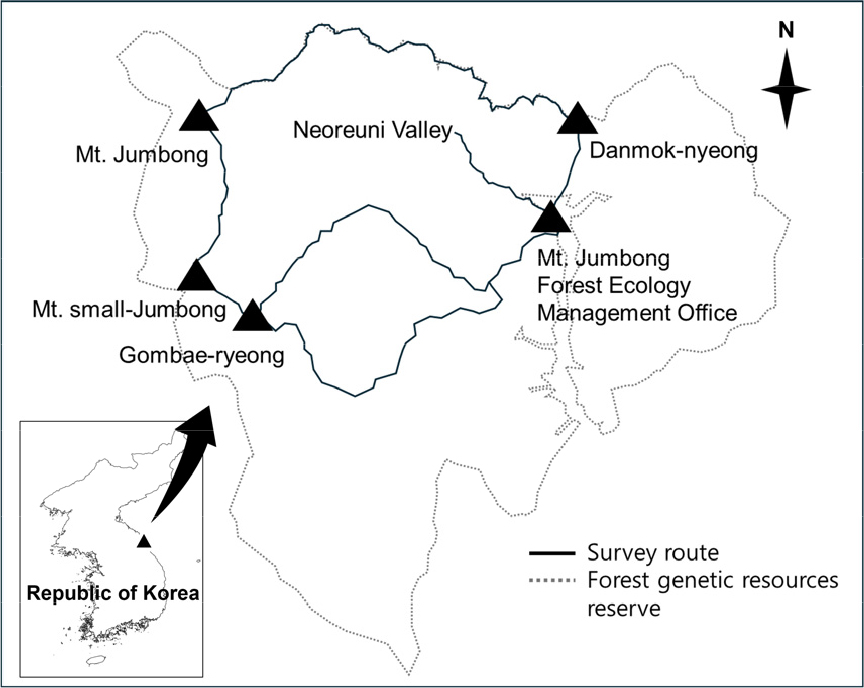
2.1 조사지역 및 시료 채집
본 연구의 조사지역인 점봉산은 1987년부터 산림유전자원보호구역으로 지정하여 보호하는 지역으로, 조사 대상 지역 약 13 km2 내에 참나무류 면적은 약 59%, 참나무류 외 기타 활엽수림이 약 30%, 침엽수림이 5% 정도로 차지하고 있으며(Korea Forest Service 2024) 관속식물류가 581분류군으로 종다양성이 매우 높은 지역이다(Korea National Arboretum 2012). 곰배령 지역은 관광객과 등산객의 이용이 빈번하긴 하나 하루 최대 등산객들의 입산이 450명으로 제한되어 있다(National Recreation Forest Management Office n.d.). 또한 곰배령 – 작은점봉산 – 점봉산 – 단목령 – 점봉산생태관리센터로 이어지는 경로는 백두대간보호지역으로 일반인들의 입산을 제한하고 있으나 백두대간 종주를 하는 일부 탐방객이 이용하고 있으며 생물다양성을 보호와 야생동물 서식지 보전을 위한 노력이 지속적으로 이루어지고 있는 지역이다(Korea National Arboretum 2012).
배설물 시료의 채집은 선조사법(Line transect sampling)을 적용하여 등산로를 따라 이동하여 주변의 고라니 배설물을 수집하였다(Fig. 1). 배설물 수집은 계절 별 먹이원의 다양성을 반영하기 위해 4월부터 10월까지 두 달 간격으로 진행되었다. 10월의 경우, 동일 조사경로상에 배설물이 확인되지 않아 이후 분석에서 제외하였다. 배설물 수집은 시료 채집시 중복 및 시료간 오염을 피하기 위하여 분변 무더기를 대상으로 3~5개 내외의 분변 펠릿을 수집하고, 수집되지 않은 펠릿은 다음 조사를 위해 흔적을 제거하였다. 채집된 배설물은 위치와 번호를 기록하고 지퍼백에 보관한 후 아이스박스에 넣어 연구실로 운반하였다. 이후 분석 전까지 –20°C 냉동고에 보관하였다.
2.2 배설물시료의 DNA 추출 및 염기서열증폭
배설물 시료에서 DNA 추출은 QIAamp® Fast DNA Stool mini kit (Qiagen, Germany)를 사용하였으며, 시료 균질화 단계를 제외하고 제조사의 프로토콜에 따라 추출하였다. 배설물 시료의 파쇄 및 균질화를 위하여 고라니의 분변시료 일부(200 mg)를 2ml Safe-Lock(R) tube (Eppendorf, Germany)에 넣고 1 ml의 Inhibit EX buffer와 5 mm 크기의 stainless steel beads를 넣어 Tissue Lyser (Qiagen, Germany)로 25 Hz에서 2분 간 파쇄를 진행하였다. 시료가 충분히 균질화 되지 않은 경우 추가로 2분간 더 파쇄를 진행하였다.
염기서열 증폭단계(polymerase chain reaction, PCR)는 고라니 배설물 내 식물성 먹이원의 DNA를 증폭하기 위해 ribosomal RNA gene의 ITS2 영역을 대상으로 하는 ITS2 S2F – S3R 프라이머 조합을 사용하였다(Table 1, Chen et al. 2010). 해당 프라이머 조합에는 이후 library 제작을 위해 Nextera transposase adaptor 서열을 부착하였으며, 시퀀싱 시 cluster generation이 원활히 진행되도록 6개의 무작위서열(N)을 배치하였다(Miya et al. 2015, illumina 2025a). PCR 혼합물은 AccuPower® HotStart premix (Bioneer, Korea)를 사용하여, 추출된 DNA의 1/10 희석액 2 µl, 프라이머(5 pmol) forward와 reverse 각 1 µl 씩 혼합하고 PCR certified water (iNtRON Biotechnology, Korea)를 이용하여 최종 20 µl 부피로 혼합하였다. PCR 조건은 초기 변성단계(initial denaturation)는 94°C에서 5분으로 설정하였고, 변성단계(denaturation)는 94°C에서 30초, 결합단계(annealing)는 56°C에서 30초, 신장단계(elongation)는 72°C에서 45초로 총 45회 진행하도록 설정하였으며 최종 신장단계(final elongation)는 72°C에서 10분 간 진행하였다. 증폭된 1차 PCR 산물은 1.5% agarose gel 상에서 증폭여부를 확인한 후 AccuPrep® PCR/Gel Purification kit (Bioneer, Korea)를 사용하여 정제되었다.
Table 1.
Primer sets used in the first PCR and the second PCR. First PCR primer sets were used to amplify DNA of plant food sources in the fecal samples. The sequences in parentheses were added to the ITS2 primers from Chen et al. (2010), representing the Nextera transposase adapter sequence and random hexamers included to improve resolution during the cluster generation step. The boldfaced sequences represent the original sequences from the cited paper. Second PCR primer sets were used for indexing, where the parentheses indicate the i7 and i5 index names, respectively
| Target | Primer | Sequence (5’ – 3’) | Reference |
| ITS2 | ITS2_S2F_modified |
(TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG NNNNNN)ATGCGATACTTGGTGTGAAT | Chen et al. 2010 |
| ITS2_S3R_modified |
(GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAG NNNNNN)GACGCTTCTCCAGACTACAAT | ||
| 1st PCR products | Index 1 Read |
CAAGCAGAAGACGGCATACGAGAT (i7-8 bp)GTCTCGTGGGCTCGG | Illumina. 2025a |
| Index 2 Read |
AATGATACGGCGACCACCGAGATCTACAC (i5-8 bp)TCGTCGGCAGCGTC |
2.3 차세대염기서열분석(Next Generation Sequencing)
본 연구에서는 illumina의 Miseq (300PE) 플랫폼에서 시퀀싱을 수행하였으며, library 제작을 위해 1차 PCR 산물에 illumina index 서열을 부착하는 2차 PCR을 진행하였다. 2차 PCR의 프라이머는 illumina PCR kit에서 제공하는 PCR 프라이머 서열을 기반으로 제작하였다(illumina 2025a, 2025b). Linker와 NexTera concensus 서열 사이에 8 bp의 i7 또는 i5의 index 서열이 존재하여 각 시료를 마킹할 수 있다(Table 1). 2차 PCR은 정제된 1차 PCR산물을 agarose gel 상에서 확인한 후 상대적인 농도에 따라 1, 2, 5, 10 µl를 주형가닥으로 사용하여, Index 1과 2 Read에 해당하는 프라이머를 각 1 µl 씩 넣어준 뒤 PCR certified water (iNtRON Biotechnology, Korea)로 총 20 µl 부피에 맞추어 진행하였다. PCR 사이클은 1차 PCR과 동일하게 진행하였으나 사이클의 반복수는 10회로 진행하였다. 증폭이 끝난 2차 PCR 산물은 Quant-ItTM PicoGreenTM dsDNA Reagent and Kit (InvitrogenTM, United States)를 이용하여 double-strand DNA의 상대농도를 측정한 후 동일한 농도로 pooling을 진행하였으며 AccuPrep® PCR/Gel Purification kit (Bioneer, Korea)를 사용하여 정제하였다.
제작한 library는 마크로젠(Macgrogen Inc., Korea)에 의뢰하여 시퀀싱과 서열 분석을 진행하였다. 서열 분석 및 분류학적 동정은 DADA2 파이프라인을 이용하여 ASV (amplicon sequence variant)를 도출한 후 NCBI (National Center for Biotechnology Information)의 nucleotide 데이터베이스를 이용하여 BLAST를 통해 수행하였다. 동정한 ASV 서열은 query coverage 95% 이상, identity percentage 97% 이상인 서열만 선별하였으며 각 샘플에서 1% 미만으로 출현한 서열은 분석에서 제외하였다(Deagle et al. 2019, Sonsthagen et al. 2020). 정리된 서열은 속 수준으로 동정하였으며, 국내 서식이 확인되지 않은 서열은 분석에서 제외하였다. 동정된 서열의 최종 명칭과 분류학적 기준은 국가생물종목록을 따라 정리하였다(National Institute of Biological Resources 2023).
2.4 데이터 분석
고라니 배설물에서 출현한 먹이원의 비율을 계산하기 위해 Deagle et al. (2019)에서 제시한 방법론에 따라 먹이원의 출현빈도(frequency of occurrence, FOO)와 서열의 상대풍부도(relative read abundance, RRA)를 계산하였다.
FOO는 먹이원의 출현 또는 비출현의 이진 데이터를 기준으로 계산되며, 먹이원 i 가 시료 k 에서 출현한 경우 li,k = 1 로 계산하고, 출현하지 않은 경우 li,k = 0 이 된다(Eq. 1). 계산식에서 S 는 총 시료의 개수를 뜻한다. RRA는 한 시료에서 발생한 모든 read 수와 해당 시료에서 발생한 먹이원 i 의 비율을 계산한 후 모든 시료에서 먹이원 i 가 차지하는 비율을 산정하여 구할 수 있다(Eq. 2). 해당 식에서 T 는 발견된 총 먹이원의 종 수를 뜻한다. 먹이원의 RRA는 속 수준에서 계산한 후 분류학적 계층에 따라 나누었고, FOO의 경우 과 별로 계산하였다. 또한 Guo and Zhang (2005)의 연구와 Lee et al. (2022)의 연구, 그리고 한반도의생물다양성(2023)의 해당 속에 대한 설명을 참고하여 검출된 식물 분류군을 속 단위에서 광엽초본(forbs), 협엽초본(graminoids), 그리고 목본(woody plants)으로 나누어 RRA를 계산하였다.
각 시료에서 발견된 먹이원 종 조성에 대한 유사도를 파악하기 위해 비계량적 다차원척도법(non-metric multidimensional scaling, NMDS)을 수행하였다. 분류군별 read counts 데이터는 속(genus)을 기준으로 log2를 취하여 정규화를 진행하여 사용하였다(Adamo et al. 2020, Skelton et al. 2023). 또한 분석 시 전체 read counts의 0.1% 이상 해당하는 속에 대해서만 진행하였다. NMDS 분석 시 read counts를 고려하기 위해 거리행렬은 Bray-Curtis dissimilarity를 선택하였다. 또한 4월과 6월 시료 간의 차이에 기여한 분류군을 선별하기 위해 동일한 처리가 된 데이터셋으로 similarity percentages (SIMPER) 분석을 수행하고 월 별 먹이원의 차이가 유의한지를 판별하기 위해 permutation analysis of variance (PERMANOVA) 분석을 수행하였다. permutation은 99,999 회로 설정하여 진행하였다. 모든 통계 분석은 통계 프로그램 R (version 4.5.0) 상에서 vegan (version 2.6-10) 패키지를 사용하여 진행하였다(Oksanen et al. 2025).
3. 결 과
3.1 섭식 먹이원 구성
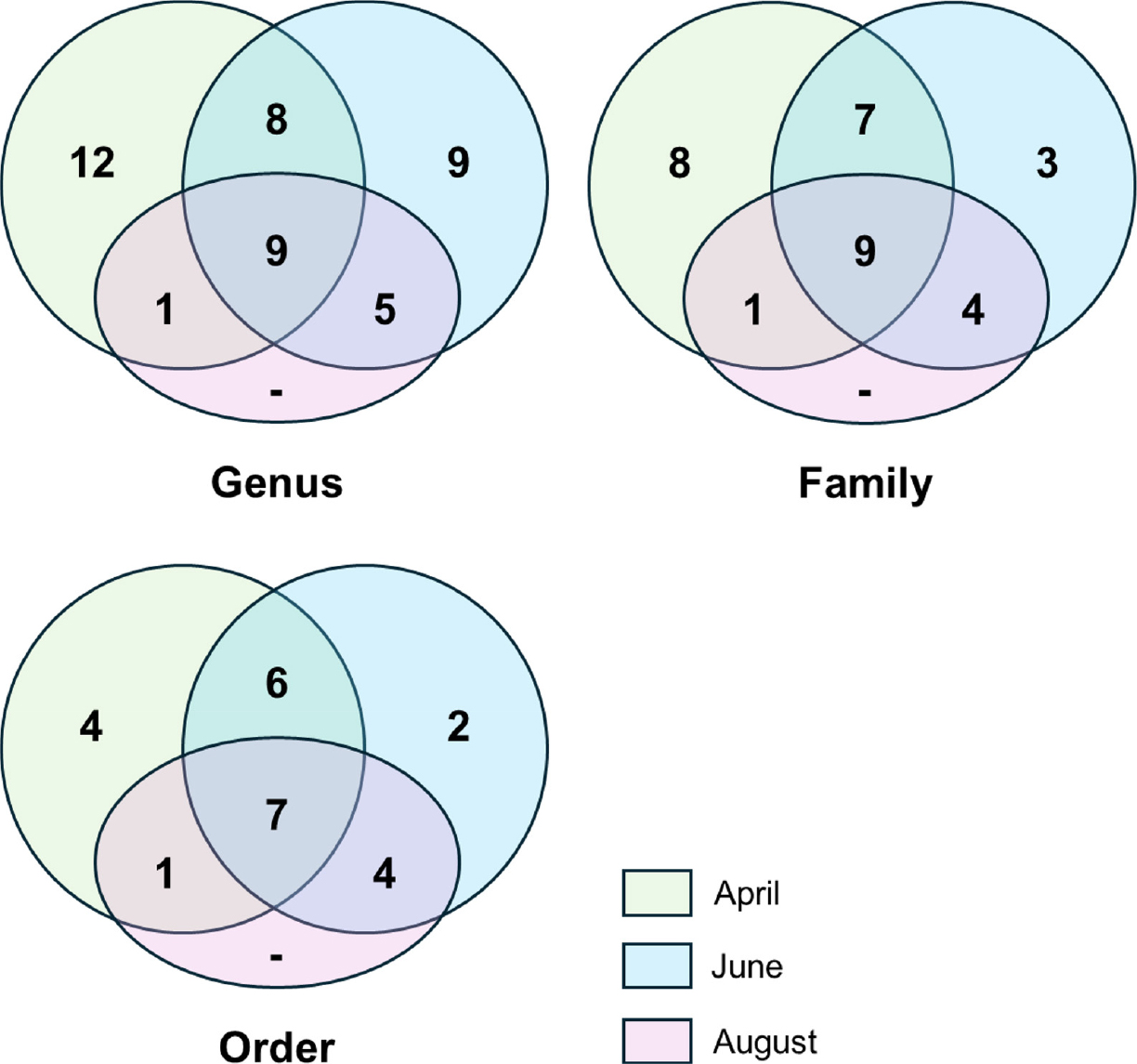
고라니 배설물은 총 4회 조사(4, 6, 8, 10월) 중 배설물이 확인되지 않은 10월을 제외하고 총 22개(4월: 9개, 6월: 11개, 8월:2개)가 채집되었다. 차세대염기서열분석(Miseq) 결과, 582개의 ASV를 확보하였으며, 분석에 사용한 유효한 read수는 총 283,362개로, 시료 당 평균 12,880.1 (± 7,853.6)의 read수를 보였다. 그 결과 총 24목, 32과, 44속의 식물을 배설물로부터 검출하였다(Table 2, Fig. 2). 전체 read counts 수가 가장 높은 속은 뽕나무속(39,840 reads)이었으며, 그 외에도 서어나무속(37,857 reads), 단풍나무속(37,137 reads), 음나무속(23,799 reads), 참나무속(19,495 reads) 등 점봉산에서 자생하는 것으로 알려진 목본식물이 다수 발견되었고 초본류 중에는 황새냉이속(17,208 reads)이 가장 높았다.
Table 2.
List of plant food sources detected in the fecal samples of Hydropotes inermis argyropus. The “Total” column indicates the number of read counts. Total relative read abundance (RRA) and frequency of occurrence (FOO) for each genus are also provided. The “Sampling month” columns show the relative read abundance (RRA) for each month
RRA 분석 결과에서는 뽕나무속, 단풍나무속, 서어나무속, 주목속, 참나무속의 RRA가 높게 나타났으며 목 수준에서는 참나무목(Fagales)의 비율이 가장 높았고 그 다음으로는 뽕나무속이 속한 쐐기풀목(Urticales), 단풍나무속이 속한 무환자나무목(Sapindales)의 비율이 높은 것으로 나타났다(Table 2, Fig. 3a). 그 외에도, 두릅나무속, 음나무속, 당귀속이 속한 미나리목(Apiales), 주목속이 속한 주목목(Taxales) 순으로 비율이 높게 나타났다. FOO 결과에서는 속 수준에서는 단풍나무속, 참나무속, 서어나무속, 소나무속, 주목속이 높게 나타났으며, 과 수준에서는 단풍나무과(Aceraceae), 자작나무과(Betulaceae), 참나무과(Fagaceae), 장미과(Rosaceae), 노박덩굴과(Celastraceae)의 빈도가 매우 높게 나타났다(Table 2, Fig. 3b). 초본류 중 FOO가 가장 높았던 분류군은 사초과(Cyperaceae)의 사초속이었다. 전체 시료 내 RRA와 FOO가 모두 높았던 분류군은 모두 목본류로 단풍나무속, 참나무속, 서어나무속, 주목속이었다. 뽕나무속의 경우 RRA 높은 편이었으나 FOO는 27.27%로 낮게 나타났으며, 반면 소나무속의 경우 전체 시료에서 RRA는 낮았으나 FOO는 비교적 높은 40.90%로 나타났다.
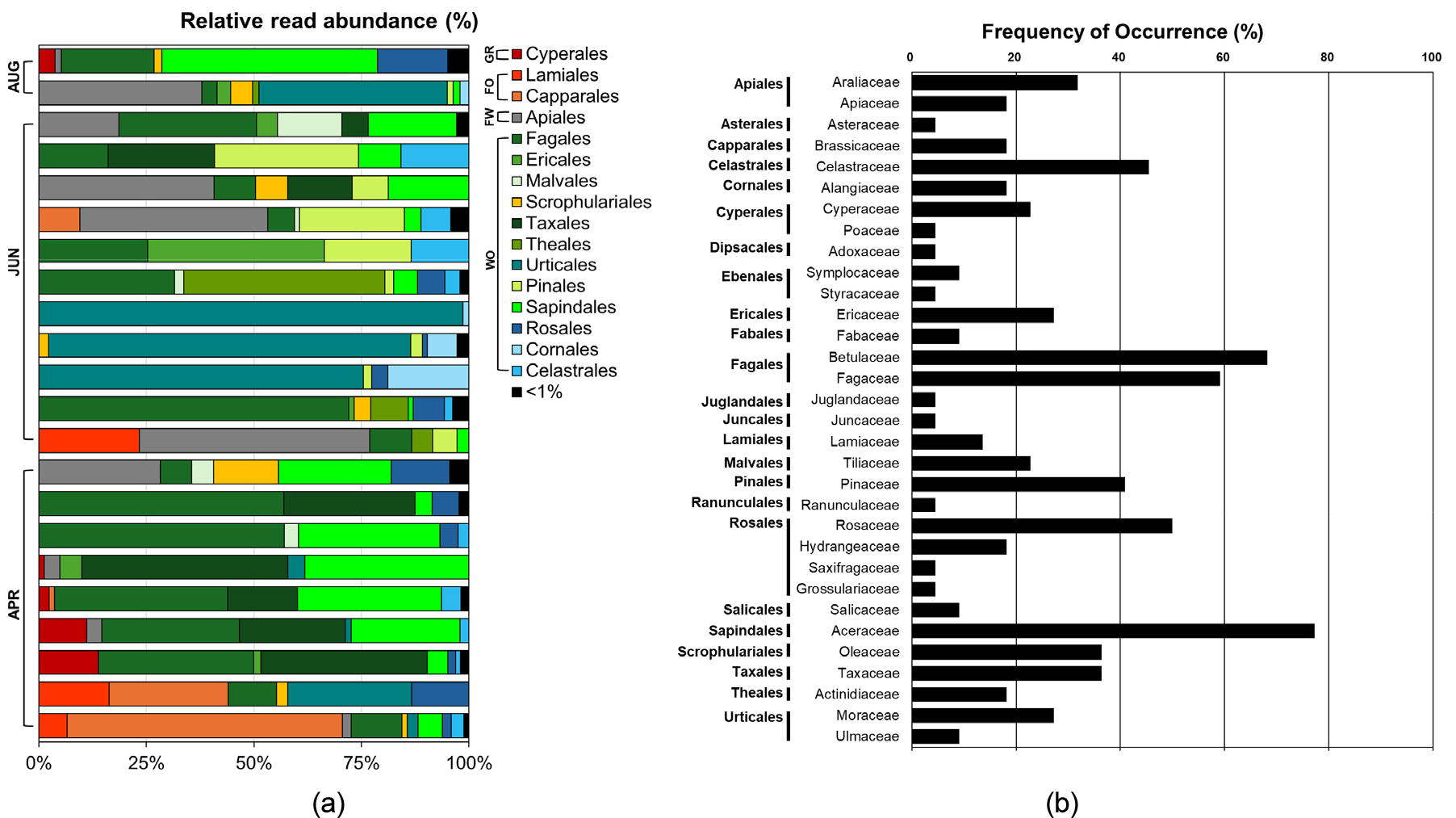
Fig. 3.
Prey items detected in the feces of Hydropotes inermis argyropus. (a) Relative read abundance of prey items from each fecal sample at the order level. Prey items with relative read abundance of less than 1% were merged and represented as “< 1%.” WO: woody plants, FW: mixture of forbs and woody plants, FO: forbs, GR: graminoids. (b) Frequency of occurrence for prey items at the family level.
3.2 먹이원의 계절별 차이
계절별 먹이원 분석은 4월과 6월 시료에 대해서 진행하였다. 계절에 따라 배설물 내에서 검출된 분류군은 차이를 보였으며 4월의 경우 총 21목 36과 51속, 6월에는 18목 28과 46속의 식물이 검출되었다(Fig. 2). 4월에는 주로 초본류(forbs)와 벼과류(graminoids)의 비율이 가장 높았으며, 6월의 경우에는 초본류 비율이 줄어든 대신 목본(woody plants)의 비율이 늘어나는 양상을 보였다(Table 3).
Table 3.
Relative read abundance (RRA) of plant food sources detected in the fecal samples of Hydropotes inermis argyropus. The symbol ‘#’ denotes the number of plant genera
| Total | APR | JUN | AUG | |||||
| # | RRA | # | RRA | # | RRA | # | RRA | |
| Forbs | 9 | 11.14 | 7 | 13.59 | 4 | 11.05 | 1 | 0.64 |
| Woody plants | 32 | 87.29 | 20 | 82.98 | 27 | 88.95 | 13 | 97.50 |
| Graminoids | 3 | 1.57 | 3 | 3.43 | - | - | 1 | 1.86 |
| Total | 44 | 100 | 30 | 100 | 31 | 100 | 15 | 100 |
먹이원의 계절간 차이를 확인하기 위해 NMDS 분석과 PERMANOVA 분석을 진행하였다. NMDS 결과에서는 4월과 6월 먹이원 군집 간의 가시적인 차이는 관찰되지 않았으나(stress = 0.155), PERMANOVA 결과에서는 통계적으로 유의한 차이를 보였다(R2 = 0.7048, F = 1.9524, p = 0.03432), (Fig. 4a). 먹이원의 월 별 차이에 기여한 분류군을 판별하기 위해 SIMPER 분석을 진행한 결과, 서어나무속(Carpinus), 소나무속(Pinus), 주목속(Taxus), 단풍나무속(Acer), 사초속(Carex), 황새냉이속(Cardamine)이 선택되었다(Fig. 4b).
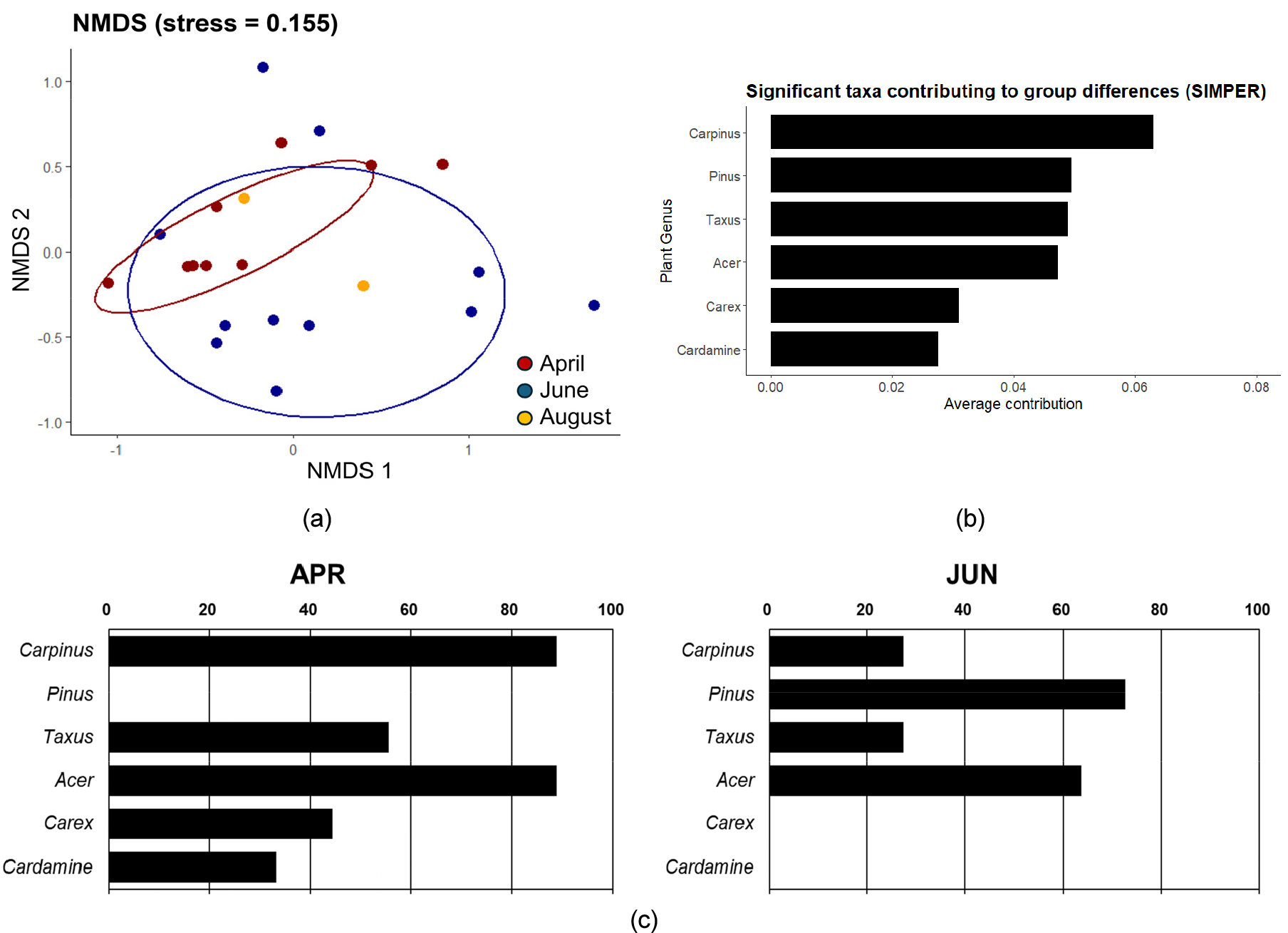
Fig. 4.
(a) Non-metric multidimensional scaling (NMDS) analysis of prey items detected in Hydropotes inermis argyropus feces by sampling month. (b) Significant plant taxa contributing to group differences between April samples and June samples, calculated by SIMPER (similarity percentages). (c) Frequency of occurrence of plants detected in fecal samples of Hydropotes inermis argyropus at the genus level. Genus selected based on SIMPER (similarity percentage) analysis.
SIMPER에서 선택된 먹이원의 차이를 확인하기 위해 월 별 RRA와 FOO를 집계하였다(Table 2, Fig. 4c, Appendix). 소나무속은 6월 샘플에서만 발견되었고, 서어나무속, 사초속, 황새냉이속은 4월 시료에서 검출 빈도가 월등히 높았다. 이 중 사초속과 황새냉이속은 4월 시료에서만 검출되었다. 단풍나무속과 주목속의 경우 FOO에는 큰 차이가 없었지만 RRA를 비교했을 때 4월 샘플에서 섭식 비율이 높은 것으로 나타났다.
4. 고 찰
야생동물의 먹이원에 대한 정보는 종의 자원 이용에 대한 행동적 특성을 설명하는 가장 큰 요인 중 하나로 서식지 보전이나 관리 전략을 위한 기초 자료로서 활용될 수 있다. 본 연구에서는 점봉산 산림 지역에 서식하는 고라니의 먹이원에 대한 정보를 제공하고, 계절적 차이를 반영하여 해당 종의 주요 먹이원을 제시하고자 하였다. 점봉산 고라니의 주된 먹이원은 목본류가 차지하고 있었으며(Table 3), 전체 시료에서 RRA와 FOO가 모두 높았던 참나무속, 단풍나무속, 서어나무속이 점봉산 고라니의 주요 먹이원으로 이용되었을 것으로 추정된다.
국내 산림지역에서 수행된 다른 선행 연구에서는 뽕나무속, 산딸기속, 벚나무속, 개암나무속, 아까시나무속의 비율이 높게 나타난 바 있다(Lee et al. 2022). 본 연구에서도 전체 시료에서 뽕나무속의 RRA가 15.15%로 가장 높게 나타났으며 산딸기속, 벚나무속의 섭식을 확인할 수 있었으나 RRA 비율은 뽕나무속, 단풍나무속, 서어나무속, 주목속 순으로 높게 나타났다(Fig. 2, Table 2). 고라니는 browser이면서 서식지 환경에서 가용한 먹이자원을 섭식한다고 알려져 있다(Hofmann 1989, Kim et al. 2021b, Kim et al. 2011a, Guo and Zhang 2005). 본 연구에서는 조사지의 식물상을 직접 관찰하지 않았으나, 점봉산 일대 주요 구성 수종에 대한 연구에서 해당 속의 식물인 신갈나무, 당단풍, 까치박달이 수평적 생태지위폭이 넓고 중층 및 하층에서의 구성 비율이 높은 종이었음을 보았을 때(Kim and Kim 2001) 점봉산 고라니의 주요 먹이원이 서식지의 식물상을 반영하였을 가능성이 있다.
고라니 먹이원의 계절적 차이를 본 연구에서 반영하고자 조사를 진행했지만 8월과 10월의 샘플 수집에 어려움이 있어 4월(봄)과 6월(초여름) 비교만이 가능하였다. 점봉산 고라니는 대부분 목본을 섭취하였으나 6월에 비해 4월의 초본류 섭식 비율이 소폭 높은 것을 확인할 수 있었으며, 선행연구와 동일하게 봄철에 초본류의 비율이 높고 겨울로 갈수록 목본의 비율이 높아질 것으로 예상되나(Guo and Zhang 2005, Kim et al. 2021b), 이와 같은 경향을 확인하기 위해서는 가을과 겨울 먹이원에 대하여 추가적인 연구가 필요하다. SIMPER 분석 결과에서는 서어나무속, 소나무속, 주목속, 단풍나무속, 사초속, 황새냉이속이 4월과 6월의 차이에 기여한 먹이원으로 선택되었다. 확인된 먹이원들이 RRA와 FOO에서도 가시적인 차이가 있었으나, 이러한 차이가 계절에 따라 가용한 먹이자원의 변화로 인한 것인지 확인하기 위해서는 가을과 겨울 먹이원을 추가로 확보할 필요가 있다. 또한 본 연구에서 사용한 DNA를 이용한 먹이원 검출 방법은 식물체의 어떤 부위를 섭식했는지에 대해서는 알 수 없다. 따라서 카메라트래핑 조사와 병행하거나, 또는 고라니 배설물 시료 채집 시기에 먹이원 식물의 성장 정도가 함께 조사된다면 먹이원의 계절적 차이가 식생의 계절적 변화를 반영하는지 더 정확하게 확인할 수 있을 것이다.
본 연구에서는 차세대염기서열분석법을 적용하여 배설물내의 DNA를 통한 비침습적인 먹이원 분석을 수행하였다. 이와 같은 배설물 내 먹이원 연구는 소화과정을 거치며 절편화된 DNA로 인하여 짧은 부위를 증폭하는 프라이머 조합을 사용하는 것이 증폭 효율이 높다고 알려져 있다(Pompanon et al. 2012). 또한 ITS2 영역은 식물종을 탐지하기 위한 메타바코딩 연구에서 주로 사용하는 영역이며, 근연종 간의 유의한 차이가 있어 짧은 길이임에도 탐지 가능한 식물성 먹이원의 범위가 넓어 많은 연구에서 사용하고 있다(Chen et al. 2010, Richardson et al. 2015, Moorhouse-Gann et al. 2018). 본 연구에서도 해당 프라이머 조합을 적용하여 44속의 다양한 식물성 먹이원을 확인하였다. 그러나 프라이머 마다 대상종에 대한 해상도 차이가 있으며, 점봉산에서의 잠재먹이원에 대한 유전자 정보(local reference database) 구축이 선행되었을 때 보다 더 정확한 해석이 가능하다(Pompanon et al. 2012, Burgar et al. 2014, Liu et al. 2021, Bänsch et al. 2020). 따라서 추후 먹이원에 대한 정확한 종 동정 및 계절별 다양한 먹이원 분석을 위해 복합적인 프라이머 조합을 이용하여 분류군별 동정 해상도를 높이거나, local database 구축을 진행한다면 더욱 정밀한 결과를 얻을 수 있을 것으로 판단된다.
5. 결 론
점봉산에 서식하는 고라니는 참나무속, 단풍나무속, 서어나무속을 주 먹이원으로 이용하며, 이외에도 뽕나무속, 주목속, 소나무속 등 목본 식물이 매우 높은 비율로 검출되었다. 이러한 결과는 고라니의 먹이원 선택에 점봉산의 식물상이 반영된 것으로 보인다. 4월과 6월 시료를 비교한 결과 4월보다 6월의 초본류 섭식 비율이 매우 낮은 것으로 확인되었으며 선행연구와 같이 겨울로 갈수록 먹이원에서 목본 비율이 높아질것으로 예상되나, 본 연구에서 가을철과 겨울철의 먹이원은 확인하지 못했기에 연중 먹이 선택에서 어떠한 경향성을 도출하기에는 한계가 있었다. 또한 서어나무속, 소나무속, 주목속, 단풍나무속, 사초속, 황새냉이속에서 계절에 따른 섭식 빈도와 비율에 차이가 확인되었다. 그러나 이러한 차이가 식생의 계절적 변동으로 인한 것인지 여부는 본 연구에서는 규명할 수 없었다. 계절에 따라 가용한 먹이 자원이 바뀌는 것으로 생각되지만 이에 대해서는 식생조사나 카메라트래핑 조사와 병행한 추가 연구가 필요할 것으로 보인다. 본 연구에서는 차세대염기서열분석법을 활용하여 비침습적인 방법으로 점봉산에 서식하는 고라니의 먹이원 분석을 수행하였으며, 향후 고라니의 관리 대책 수립 및 보전 정책 마련을 위한 기초 생태학적 자료를 제공하였다.
