ⓒ Korean Society of Ecology and Infrastructure Engineering. All rights reserved.
ABSTRACT
1. 서 론
2. 연구 방법
2.1 조사지 개황
2.2 현장 조사
2.3 자료 처리
3. 결과 및 고찰
3.1 식물상 특성
3.2 식생 구조 특성
4. 하천변 침입외래식물 관리를 위한 제안
1. 서 론
하안역 (riparian zone)은 하천을 따라서 형성된 계절적 또는 간헐적으로 범람하는 지역이다. 하안역은 홍수 저감, 수질 정화, 퇴적 완충, 지하수 재충전, 야생생물 서식지로서 기능을 제공한다 (Naiman and Décamps 1997). 이곳에 분포하는 하안 식생은 종횡적 하천 환경의 이질성에 의하여 생물다양성이 높고 홍수 교란에 의존적인 초기 천이단계의 특성을 보인다 (Masters and Sheley 2001). 하안역은 인간 활동과 관리자 활동이 집중되는 장소로서 댐 축조와 물흐름 조절에 의한 수문 변경, 하천변 토지 이용, 벌목, 답압 및 휴양 등의 다양한 인위적 교란이 일어나고 있다 (Richardson et al. 2007). 이러한 자연적, 인위적 교란은 전형적인 천이초기종인 외래 식물종의 침입과 확산을 촉발 또는 촉진할 수 있다. 따라서 하안역을 침입종의 안식처로서 비유하기도 한다 (Stohlgren et al. 1998).
하안역에서 침입외래종 (invasive alien species)은 생물계절, 형태 및 생태적 지위 요구가 고유종과 다르므로 물리적 및 생물적 환경과 영향을 다르게 주고 받을 것으로 생각할 수 있다. 따라서 침입외래종이 번성하는 하안역은 고유의 전형적인 구조와 기능에서 벗어난 특성을 보이게 된다. 무엇보다 먼저 침입외래종에 의하여 하안 고유 생물군집의 구조가 변화되는데 특히 하천 고유 개척자 식물종이 피해를 받아서 생물다양성이 변형된다 (Shin and Cho 2001, Asaeda et al. 2011). 이러한 식물군집의 조성과 구조의 변화는 생태계 기능을 다양하게 변화시킬 수 있다 (Richardson et al. 2007). 예를 들면 침입식물의 생물량 증가는 식생에 의한 물 흡수가 증가되어 하천유량을 감소시키거나 (Van Lill et al. 1980), 수리 조도가 증가되어 유사가 퇴적되고 하도의 하상 상승을 유발시킬 수 있다 (Zavaleta et al. 2001).
우리나라의 환경부에서는 침입외래식물 중에서 고유생태계에 심각하게 부정적인 영향을 미치는 12종을 생태계교란생물로서 지정하여 특별 관리하고 있다 (NIER 2012). 이중에서 하천 수변부에서 생태적, 관리상의 문제를 유발하고 있는 대표적인 침입외래식물이 가시박과 털물참새피이다 (NIE 2014). 가시박 (Sicyos angulatus)은 자생지가 북미 중동부이고 (USDA 2015), 우리나라를 비롯한 동아시아, 멕시코, 중미, 유럽에 도입되어 있다 (EPPO 2010). 이 종은 박과의 일년생 식물로서 덩굴 줄기가 매우 빠르게 성장하여 농작물이나 자연식생을 감고 덮어서 큰 문제를 일으키고 있다. 습지지수 (wetland status)는 임의 (facultative) 또는 임의습지 (facultative wetland)이다 (USDA 2015). 또 다른 교란식물인 털물참새피 (Paspalum distichum var. indutum)는 원산지가 북미 동부 및 남부이지만 (USDA 2015), 현재 세계 각지로 도입되어 우리나라를 비롯한 동아시아에서부터 열대 아메리카까지로 폭 넓게 분포하고 있다 (NIER 2012). 털물참새피는 벼과에 속하는 다년생 초본으로서 지면이나 수면을 기거나 뜨는 줄기를 뻗어서 수면을 완전히 덮는 단순 군락을 형성하여 다른 식물종과 어류의 생존을 위협하여 생물종다양성을 감소시키는 부정적인 영향을 일으키고 있다 (Yang et al. 2002, Stroh 2006). 이 식물의 습지지수는 임의습지 (facultative wetland) 또는 절대습지 (obligate wetland)이다 (USDA 2015).
본 연구에서는 전국을 대상으로 가시박과 털물참새피가 침입한 하천 수변부에서 식생 구조를 조사하여 이들 침입외래식물에 의하여 변형된 수변부 식생의 구조적 특성을 파악하고 이 결과를 바탕으로 합리적 식생 관리 방안을 제안하고자 하였다.
2. 연구 방법
2.1 조사지 개황
우리나라 하천에 분포하는 침입외래식물종으로서 환경부 지정 생태계교란생물인 가시박 (Sicyos angulatus)과 털물참새피 (Paspalum distichum var. indutum)가 우점하는 식물군집의 식물상과 식생구조를 전국적으로 각각 11지소를 선정하여 조사하였다 (Fig. 1 및 Table 1). 선정된 조사지점은 국립생태원이 수행하는 생태계교란종 모니터링 조사지에 포함된다 (NEI 2014). 가시박 조사지점은 이들이 주로 분포하는 남한의 중북부 지역에 한정되어 있었다. 가시박이 출현하는 지역은 주로 하천 고수부, 제방사면, 도로변 등인데 이중에서 하천 고수부, 제방 사면 및 상부를 포함하는 하천변에서 조사지점을 선정하였다. 이중에서 조사지점 S5는 강원도 춘천시 의암호의 호변에 위치한 곳이나 이곳 또한 하천변과 유사한 환경으로서 본 연구의 조사지점에 포함하였다. 각 조사지점에서 가시박은 교관목이나 장경초본을 덩굴로서 완전히 덮고 있었다 (Fig. 2). 털물참새피 조사지점은 이들이 주로 분포하는 전라북도 정읍시 (Fig. 1의 P1) 이남의 전라남도와 경상남도의 한반도 남부지역에서 11 곳을 선정하였다 (Fig. 1 및 Table 1). 털물참새피의 서식지 유형은 유입하천을 포함하는 저수지 수변과 하천의 유속이 느린 저수로 또는 홍수터 습지의 물가에서 물 위를 덮고 있었다 (Fig. 2).
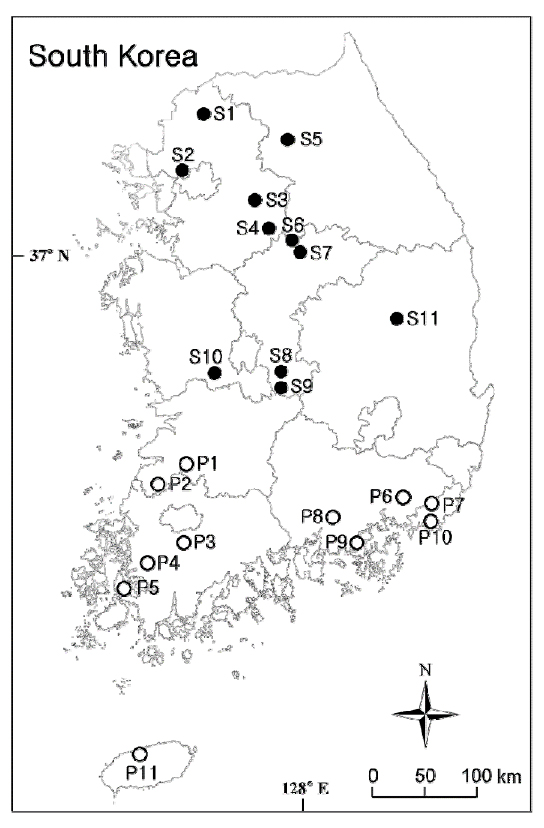
|
Fig. 1. Map showing the sampling sites of Sicyos angulatus (the closed circles) and Paspalum distichum var. indutum (the open circles). |

|
Fig. 2. Photographs showing the typical habitats of Sicyos angulatus (a) and Paspalum distichum var. indutum (b). |
Table 1. Location and habitat type of sampling sites for Sicyos angulatus and Paspalum distichum var. indutum. |
Site no. | Address | Latitude | Longitude | Habitat |
Sicyos angulatus |
S1 | Jeongok-eup, Yeoncheon-gun, Gyeonggi-do | 38° 00′ 20.3″ N | 127° 03′ 29.8″ E | Streamside |
S2 | Daeja-dong, Deogyang-gu, Goyang-si, Gyeonggi-do | 37° 41′ 14.4″ N | 126° 52′ 32.1″ E | Streamside |
S3 | Yangpyeong-eup, Yangpyeong-gun, Gyeonggi-do | 37° 27′ 55.0″ N | 127° 31′ 29.0″ E | Streamside |
S4 | Hadong, Yeoju-si, Gyeonggi-do | 37° 18′ 37.7″ N | 127° 37′ 02.3″ E | Streamside |
S5 | Seo-myeon, Chuncheon-si, Gangwon-do | 37° 53′ 17.7″ N | 127° 41′ 17.4″ E | Streamside |
S6 | Buron-myeon, Wonju-si, Gangwon-do | 37° 09′ 50.9″ N | 127° 45′ 55.8″ E | Streamside |
S7 | Sotae-myeon, Chungju-si, Chungcheongbuk-do | 37° 05′ 09.1″ N | 127° 50′ 30.7″ E | Streamside |
S8 | Iwon-myeon, Okcheon-gun, Chungcheongbuk-do | 36° 13′ 36.9″ N | 127° 40′ 12.3″ E | Streamside |
S9 | Gwangseok-myeon, Nonsan-si, Chungcheongnam-do | 36° 12′ 37.2″ N | 127° 04′ 41.0″ E | Streamside |
S10 | Yangsan-myeon, Yeongdong-gun, Chungcheongbuk-do | 36° 07′ 34.0″ N | 127° 38′ 26.0″ E | Streamside |
S11 | Seohu-myeon, Andong-si, Gyeongsangbuk-do | 36° 38′ 19.0″ N | 128° 41′ 48.7″ E | Streamside |
Paspalum distichum var. indutum |
P1 | Soseong-myeon, Jeongeup-si, Jeollabuk-do | 35° 34′ 34.1″ N | 126° 48′ 10.8″ E | Reservoir |
P2 | Haeri-myeon, Gochang-gun, Jeollabuk-do | 35° 28′ 24.5″ N | 126° 31′ 43.9″ E | Reservoir |
P3 | Sanpo-myeon, Naju-si, Jeollanam-do | 35° 01′ 15.6″ N | 126° 49′ 32.0″ E | Reservoir |
P4 | Samhyang-eup, Muan-gun, Jeollanam-do | 34° 50′ 48.9″ N | 126° 25′ 35.6″ E | Reservoir |
P5 | Hwawon-myeon, Haenam-gun, Jeollanam-do | 34° 38′ 42.8″ N | 126° 18′ 55.3″ E | Reservoir |
P6 | Daesan-myeon, Uichang-gu, Changwon-si, Gyeongsangnam-do | 35° 19′ 10.3″ N | 128° 41′ 02.5″ E | Stream |
P7 | Daedong-myeon, Gimhae-si, Gyeongsangnam-do | 35° 14′ 54.0″ N | 128° 59′ 15.0″ E | Stream |
P8 | Sangpyeong-dong, Jinju-si, Gyeongsangnam-do | 35° 10′ 05.2″ N | 128° 06′ 29.2″ E | Stream |
P9 | Goseong-eup, Goseong-gun, Gyeongsangnam-do | 34° 59′ 07.8″ N | 128° 19′ 55.1″ E | Stream |
P10 | Samrak-dong, Sasang-gu, Busan | 35° 09′ 05.2″ N | 128° 58′ 07.2″ E | Stream |
P11 | Naedo-dong, Jeju-si, Jeju-do | 33° 29′ 29.9″ N | 126° 26′ 32.8″ E | Stream |
2.2 현장 조사
선정된 조사지점에서 식물상과 식생구조를 2014년 4월과 5월에 조사하였고, 7월부터 9월 사이에 식물상을 보완 조사하였다. 식물상 조사는 조사지점을 직접 답사하면서 출현식물을 기록하였다. 현장에서 동정이 어려운 식물종은 식물체를 채집하여 실험실에서 동정하였다. 식물종의 동정은 Lee (2003)와 Park (2009)을 참조하였다. 동정된 식물의 종명과 외래종, 생활사. 목본 및 초본, 덩굴식물 등의 특성 분류는 KNA (2015)를 따랐다. 또한 출현식물의 습지 지표종 범주 (wetland indicator category)를 Choung et al. (2012)에 따라서 절대습지 (obligate wetland), 임의습지 (facultative wetland), 임의 (facultative), 임의육상 (facultative upland) 및 절대육상 (obligate upland) 식물로 분류하였다.
각 조사지점에서 식물군집 구조를 파악하기 위하여 각 조사지점에서 방형구를 설치하고 출현종과 종별 피도를 조사하였다. 방형구는 털물참새피 조사지점에서는 5 m x 5 m 크기로, 가시박 조사지점에서는 가시박에 의하여 피압된 식물의 높이에 따라서 1 m x 1 m, 2 m x 2 m 또는 5 m x 5 m 크기로 조사지점마다 5개를 설치하였다.
2.3 자료 처리
현장에서 조사한 식물상 자료로부터 총출현종수와 총종수에 대한 절대습지식물, 임의습지식물, 외래종, 1·2년생, 목본 및 덩굴 종수의 백분율을 구하였다. 식물상에 의한 조사지점 사이의 관계를 파악하기 위하여 주좌표분석 (principal coordinates analysis, PCoA)을 실시하였다. 조사지점별 식물상 자료에서 3 조사지점 이상에서 출현한 종의 출현 자료로서 입력 자료를 구축하였다. 정리된 입력 자료를 이용하여 R 환경 (R Development Core Team 2013)에서 Vegan package (Oksanen 2013)을 이용하여 ‘rda’ 함수로서 Bray-Curtis 거리 지수 (Bray and Curtis 1957)를 계산하고 ‘cmdscale’ 함수로서 주좌표분석을 실시하였다.
조사지점의 식생구조를 파악하기 위하여 종별 피도로 구성된 식생 자료을 이용하여 종다양도 지수를 구하고 탈경향 대응 분석 (detrended correspondence analysis, DCA)을 실시하였다. 종다양도 지수는 종별 피도 자료로부터 자연 로그를 이용하는 Shannon- Wiever 다양도 지수로 계산하였다 (Shannon and Weaver 1949). 조사지점 사이의 식생구조를 비교하기 위하여 R 환경 (R Development Core Team 2013)에서 Vegan package (Oksanen 2013)의 ‘decorana’ 함수를 이용하여 탈경향 대응 분석을 수행하였다. 2개 조사지점 이상에서 출현한 식물종의 피도 (S)를 log (S+1)로 전환하여 탈경향 대응 분석을 위한 입력 자료를 구축하였다. 한편, 조사지점의 식물상 특성과 식생구조의 관계를 분석하기 위하여, 각 조사지점별로 식물상으로부터 산출한 총출현종수와 총종수에 대한 절대습지식물, 임의습지식물, 외래종, 1·2년생, 목본 및 덩굴 종수의 백분율 및 식생구조에서부터 산출한 다양도 지수를 입력자료로 하여 Vegan package (Oksanen 2013)의 함수 ‘envfit’를 사용하여 유의한 요인을 탈경향 대응 분석 결과의 좌표 그래프에 화살표로 도시하였다.
3. 결과 및 고찰
3.1 식물상 특성
하천 제외지에 도입되어 생태적, 경관적 문제를 야기하고 있는 가시박과 털물참새피 조사지의 식물상 특성을 주좌표 분석 (PCoA)으로 파악하였다 (Fig. 3). 식물종의 배열에서 외래종 분포지에 공통적으로 나타나는 출현빈도가 높은 식물종은 그래프의 좌측 중앙에 집중되었다 (Fig. 3 a). 이들 종은 환삼덩굴 (H.ja), 쑥 (A.pr), 망초 (C.ca), 갈대 (P.cm), 왕고들빼기 (L.in), 개망초 (E.an), 소리쟁이 (R.ci), 쇠뜨기 (E.ar), 강아지풀 (S.vi), 닭의장풀 (C.co), 미국가막사리 (B.fr) 등으로서 대부분 교란지식물 (ruderals)이었다. 그래프의 좌측에 위치한 가시박 (S.an)과 가까이 위치한 종은 칡 (P.lb), 뽕나무 (M.al), 조팝나무 (S.pr) 등으로서 주로 제방 사면에 위치한 목본식물이었다. 또한 그래프 우측 중앙에 위치한 털물참새피 (P.ds)와 가까이 위치한 종은 노랑꽃창포 (I.pe), 애기부들 (T.an), 개구리밥 (S.po) 등과 같이 저수로 수변에 분포하는 식물이었다.
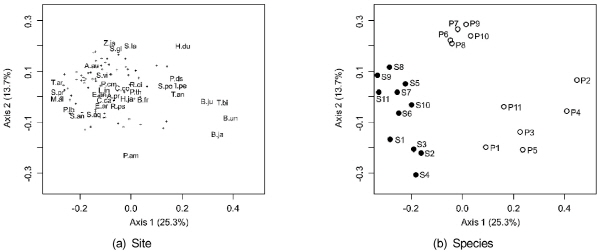
|
Fig. 3. Ordination biplots of plant species (a) and sampling sites (b) from the results of principal coordinates analysis using floristic data at the sampling sites of Sicyos angulatus (the closed circles) and Paspalum distichum var. indutum (the open circles). The abbreviations of sampling sites refer to Fig. 1. (A.au, Acalypha australis; A.pr, Artemisia princeps; B.fr, Bidens frondosa; B.ja, Bromus japonicas; B.ju, Brassica juncea var. integrifolia; B.un, Bromus unioloides; C.ca, Conyza canadensi; C.co, Commelina communis; E.an, Erigeron annuus; E.ar, Equisetum arvense; H.du, Hydrocharis dubia; H.ja, Humulus japonicus; I.pe, Iris pseudoacorus; L.in, Lactuca indica var. laciniata; M.al, Morus alba; P.am, Phytolacca Americana; P.cm, Phragmites communis; P.ds, Paspalum distichum var. indutum; P.la, Persicaria lapathifolia var. lapathifolia; P.th, Polygonum thunbergi; R.ci, Rumex crispus; R.ps, Robinia pseudo-acacia; S.an, Sicyos angulatus; S.gl, Setaria glauca; S.la, Scirpus lacustris var. crebe; S.po, Spirodela polyrhiza; S.pr, Spiraea prunifolia for. simpliciflora; S.vi, Setaria viridis; T.an, Typha angustifolia; T.ar, Thlaspi arvense; T.bi, Trapa bispinosa var. inumai; Z.ja, Zoysia japonica). |
조사지 식물상에 의한 조사지점 배열에서 가시박과 털물참새피의 조사지점이 뚜렷이 구분되었다 (Fig. 3 b). 식물종의 배열에서와 같이 가시박은 목본식물이 많은 곳에 털물참새피는 수생식물이 많은 곳에 위치하였다. 한편 그래프의 좌측에 위치한 가시박 조사지점 중에서 경기도 위치한 조사지점 (S1 – S4)과 나머지 강원도와 충청도 이남 조사지점 (S5 – S11)이 멀리 위치하였다. 경기도 조사지점에서는 가시박이 주로 하천 고수부지에 분포하였고 나머지 조사지점에서는 제방 사면과 상부에 분포하였다. 털물참새피 조사지점은 전라도와 제주도 조사지점 (P1 – P5, P11)과 경상도 조사지점 (P6 – P10)으로 구분되었다. 전라도 조사지점은 주로 저수지 수변부의 정체수역으로서 마름 (T.bi), 큰이삭풀 (B.un) 등이 특이적으로 출현하였고, 경상도 조사지점은 주로 유속이 느린 하천 수변부로서 큰고랭이 (S.la), 자라풀 (H.du) 등이 분포하였다.
식물종 특성 (traits)에 의한 식물상의 구조적 특성을 살펴보면, 임의습지 지표식물 (FACW)과 외래종의 총종수에 백분율은 가시박과 털물참새피 군집에서 유사하였다 (Table 2). 절대습지 식물 (OBW)의 백분율은 털물참새피 군집이 가시박 군집보다 높았으나, 1·2년생, 목본 및 덩굴 식물의 백분율은 가시박 군집이 털물참새피 군집보다 높았다. 또한 조사지점별 총출현종수는 가시박이 평균 50 종류로서 털물참새피의 35 종류보다 많았다 (Table 3).
Table 2. Mean, standard deviation (SD), minimum (Min) and maximum (Max) of the proportion of the number of species according to the groups of obligate wetland indicators (OBW), facultative wetland indicators (FACW), exotic plants, annuals and biennials, trees and vines at the communities of Sicyos angulatus and Paspalum distichum var. indutum (n=11). |
Group | Sicyos angulatus | Paspalum distichum var. indutum |
Mean | SD | Min | Max | Mean | SD | Min | Max |
OBW (%) | 3.6 | 2.3 | 0.0 | 8.1 | 19.5 | 13.7 | 9.7 | 57.9 |
FACW (%) | 10.9 | 6.2 | 0.0 | 21.6 | 11.2 | 5.4 | 0.0 | 17.9 |
Exotic species (%) | 26.6 | 2.6 | 22.2 | 31.5 | 24.4 | 9.9 | 0.0 | 40.0 |
Annuals & biennials (%) | 55.7 | 18.4 | 21.3 | 76.6 | 40.8 | 16.9 | 10.6 | 66.0 |
Tree (%) | 18.6 | 11.2 | 0.0 | 36.2 | 7.0 | 5.1 | 2.1 | 19.1 |
Vine (%) | 16.1 | 6.0 | 8.5 | 25.5 | 6.4 | 3.0 | 2.1 | 12.8 |
Table 3. Mean, standard deviation (SD), minimum (Min) and maximum (Max) of the number of total species and diversity index at the communities of Sicyos angulatus and Paspalum distichum var. indutum (n=11). |
Property | Sicyos angulatus (n=9) | Paspalum distichum var. indutum (n=6) |
Mean | SD | Min | Max | Mean | SD | Min | Max |
No. of species | 50 | 35 | 18 | 73 | 35 | 9 | 19 | 48 |
Diversity index | 2.05 | 0.41 | 1.56 | 2.58 | 1.14 | 0.63 | 0 | 1.75 |
3.2 식생 구조 특성
가시박과 털물참새피의 식생자료를 이용하여 탈경향 대응 분석 (DCA)를 실시한 결과에 따르면 제1축과 제2축의 고유값 (eigen value) 백분율이 각각 55.5%와 21.2%로서 두 축으로 전체 변이의 76.7%를 설명하였다 (Fig. 4). 먼저 식물종 배열을 살펴보면 털물참새피 (P.da)와 이와 동반하는 줄 (Z.la), 마름 (T.bi) 등은 그래프의 우측에 배열되었다 (Fig. 4 a). 한편 그래프의 좌측에는 가시박 (S.an)과 함께 칡 (P.lb), 족제비싸리 (A.fr), 개망초 (E.an) 등이 가까이 위치하였다.
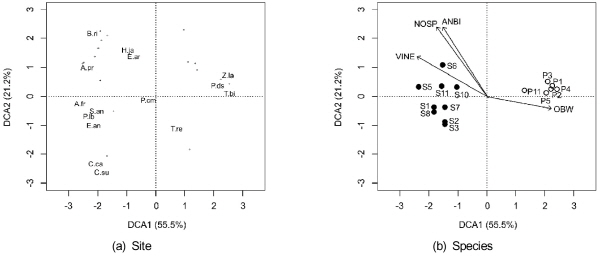
|
Fig. 4. Biplots of plant species (a) and sampling sites (b) by detrended correspondence analysis using vegetation data at the sampling sites of Sicyos angulatus (the closed circles) and Paspalum distichum var. indutum (the open circles). The angles and lengths of the radiating arrows indicate the direction and strength of relationships of total number of species (NOSP) and the proportions of plant species according to the groups of annuals and biennials (ANBI), obligate wetland indicators (OBW) and vines (VINE). The abbreviations of sampling sites refer to Fig. 1. (A.fr, Amorpha fruticosa; A.pr, Artemisia princeps, B.ri, Bromus rigidus; C.ca, Conyza canadensi; C.su, Conyza sumatrensis; E.an, Erigeron annuus; E.ar, Equisetum arvense; H.ja, Humulus japonicus; P.cm, Phragmites communis; P.ds, Paspalum distichum var. indutum; P.lb, Pueraria lobata; S.an, Sicyos angulatus; T.bi, Trapa bispinosa var. inumai; T.re, Trifolium repens; Z.la, Zizania latifolia). |
식생 자료를 이용한 탈경향 대응 분석에 의한 조사지점의 배열에서 가시박 조사지점은 제1축의 왼쪽에 털물참새피 조사지점은 오른쪽에 위치하였다 (Fig. 4 b). 특히 털물참새피 조사지점은 서로 가까이 밀집하여 분포하였으나 가시박 조사지점은 분산되어 있어서 가시박의 조사지점간 식생구조의 차이가 크다는 것을 알 수 있었다. 종다양성, 식물상 특성과 식생구조의 상관 분석에서 절대습지 (Fig. 4 b의 화살표에서 OBW), 1·2년생 (ANBI) 및 덩굴 (VINE) 식물종수 백분율과 총종수 (NOSP)가 유의한 요인으로 선정되었다 (Fig. 4). 털물참새피 조사지점이 있는 방향으로 절대습지지표 식물종 백분율이 높아서 물가에 가까운 생육지 환경임을 나타내었다. 또한 가시박이 있는 오른쪽 윗쪽으로 갈수록 총종수, 덩굴식물 및 1·2년생종 백분율이 높아서 인위적인 교란이 가해진 곳 임을 알 수 있었다. 종다양성 역시 가시박 군집이 털물참새피 군집보다 높았다 (Table 3).
이상의 결과를 종합하면 생태계교란생물로 지정된 가시박과 털물참새피는 하천 및 저수지 수변부의 교란지에서 주로 분포하였다. 이중에서 가시박은 고수부 상부와 제방사면의 목본식물이 분포하는 곳의 보다 다양한 환경에 침입하여 있었다. 반면에 털물참새피는 보다 습한 생육지를 선호하며 주로 줄, 마름 등과 같은 절대습지식물과 생태적 지위가 겹치는 것으로 생각된다. 가시박은 일본 Tama 강의 범람원에서 주로 물억새와 동반 출현하였고, 가시박이 물억새 군집에 침입하고 홍수 교란이 가해짐에 따라서 장경의 물억새 군집이 단경 초지로 변하였다 (Uchida et al. 2012). 그러나 우리나라에서는 가시박이 주로 교관목의 목본식물 군집에 침입하고 이들의 생육을 억제하는 것으로 나타났다. 일반적으로 가시박은 미립 토양, 수분, 영양소 요구가 높고 천근성이기 때문에 이 식물이 미립 하상에 침입하면 미립토의 침식 (washing)이 감소된다 (Asaeda et al. 2011). 또 다른 침입외래종인 털물참새피는 주로 유속이 느린 저수로 수변에서 수면을 덮어서 수생식물의 종다양성을 침해하고 있다. 포르투갈과 스페인의 하천에서 털물참새피는 제방사면, 홍수터 삼림보다는 주로 교란이 가해진 단절된 홍수터의 가장자리와 사주의 물가를 따라서 분포하였다 (Aguiar et al. 2005, González et al. 2015). 하천변에서 이 식물이 침입하여 물 위를 덮으면 다른 식물종과 어류의 생존을 위협하여 생물종다양성을 감소시킨다 (Yang et al. 2002, Stroh 2006). 본 연구에서도 가시박이 침입한 식생보다 털물참새피 침입 식생에서 출현 식물의 종다양성이 낮았다 (Table 3).
4. 하천변 침입외래식물 관리를 위한 제안
하천변 생태계의 보전과 복원을 위한 침입외래식물의 관리는 크게 직접 및 간접 방제법으로 구분할 수 있다 (González et al. 2015). 이중에서 직접 방제는 식물체를 직접 제거하는 기계적 방제, 제초제를 이용하는 화학적 방제, 초식자나 기생자를 이용하는 생물학적 방제를 포함한다. 또한 간접 방제는 고유종에 적합한 수리적 및 지형적 변화를 유도하거나 고유종을 직접 도입하여 외래종에 대한 고유종의 경쟁력을 강화시키는 방법이다. 우리나라 하천에서 생태적, 관리적 문제를 유발하고 있는 가시박과 털물참새피의 관리 및 방제 방안에 대하여 본 연구 결과를 바탕으로 다음과 같이 제안한다.
가시박은 현재 하천의 홍수터 상부와 제방 사면의 수목이나 물억새 등의 고경 초목 등의 다양한 생육지에서 자연 식생을 피압하고 있다. 더욱이 이 식물이 하천을 따라서 종적으로 확산되어, 침입한 하천 제외지로부터 제내지의 도로, 경작지 및 숲 가장가지까지 횡적으로도 확산되고 있는 실정이다. 가시박을 직접 제거하고자 할 때는 종자를 맺기 전에 지면에서 줄기를 잘라주면 효과적이다. 그러나 가시박의 제거가 다른 침입식물의 정착을 유도하는 이차천이를 진행시킬 수 있기 때문에 주의가 요구된다. 일본의 경우 가시박을 인위적으로 제거한 직후에 환삼덩굴, 호장근, 갓, 쥐보리 등 다른 침입식물의 정착이 진행되었다 (Hashimoto 2010). 따라서 물리적으로 가시박을 제거한 지역에 신속하게 정착할 수 있는 대체 식재종 개발의 필요성을 강조하고 있다. 간접 방제법으로서, 가시박이 하천 고수부지와 제방 사면의 양지이면서 적습한 장소를 선호하므로 지형을 낮게 유지하고 침식을 유도하여 침입지역에서 침수 또는 과습 기간을 증가시킴으로서 가시박의 생장을 억제할 수 있다. 그러나 무엇보다 중요한 것은 제외지 홍수터와 제방 사면에서 가시박 도입을 유도하는 인위적인 교란을 억제하는 것이 중요하다 (Liendoa et al. 2015).
또 다른 하천의 주요 침입식물인 털물참새피는 흐름이 정체된 곳이나 느린 곳에서 물 위를 떠서 퍼지는 게릴라 확산이 가능하여 다른 식물의 정착을 억제하고 물 속으로 투광을 억제하고 있다. 이 식물은 물 위 뿐만 아니라 땅 위를 기어서 확산할 수 있기 때문에 수위변동에 강한 특성을 지니고 있다. 또한 털물참새피는 종자, 지하경, 포복경의 다양한 방법으로 번식이 가능하여 쉽게 수면을 덮을 수 있다 (Noda and Obayashi 1971). 털물참새피는 지하경, 포복경으로 넓은 수면과 지면을 덮고 있기 때문에 기계적으로 제거하기가 쉽지 않다. 그러나 이 식물이 피음에 민감하기 때문에 (Manuel and Mercado 1977), 고마리, 미나리 등과 같은 단경 초본보다 경쟁에서 유리하지만 애기부들, 갈대 등과 같은 장경 초본보다는 경쟁력이 낮다. 따라서 유속이 느린 하천의 저수로변에 장경 초본의 생육을 촉진함으로써 털물참새피의 침입을 억제할 수 있다. 또한 털물참새피는 전식물체가 동시에 침수되는 것에 취약 하므로 (Hsiao and Huang 1989), 저수로변에서 일정 기간 수위를 상승시킴으로써 이들의 방제가 가능할 수 있다. 한편 털물참새피를 방제하면 질병 유발 모기 발생을 억제하는 부수적 이점도 얻을 수 있다. 털물참새피가 물 위를 덮은 곳에서는 모기 유충이 포식자로부터 은신처를 제공 받을 수 있기 때문에 이 식물이 침입한 수변에서 모기 발생이 증가하였다 (Greenway et al. 2003).
Acknowledgements
본 연구의 자료 분석과 논문 작성을 도와준 인하대학교 조강현 교수에게 감사드린다.
References
Aguiar, F.C., Ferreira, M.T., Albuquerque, A. and Bernez, I. 2005. Invasibility patterns of knotgrass (Paspalum distichum) in Portuguese riparian habitats. Weed Technology 19: 509-516.
Asaeda, T., Rashid, M.H., Kotagiri, S. and Uchida, T. 2011. The role of soil characteristics in the succession of two herbaceous lianas in a modified river floodplain. River Research and Applications 27: 591-601.
Bray, J.R. and Curtis, J.T. 1957. An ordination of upland forest communities of southern Wisconsin. Ecological Monographs 27: 325-349.
Choung, Y.S., Lee, Y.T., Cho, K.H., Joo, K.Y., Min, B.M., Hyun, J.O. and Lee, K.S. 2012. Categorizing Vascular Plant Species Occurring in Wetland Ecosystems of the Korean Peninsula. Center for Aquatic Ecosystem Restoration, Chuncheon, Korea. (in Korean)
EPPO. 2010. EPPO data sheet on invasive alien plants: Sicyos angulatus. European and Medi-terranean Plant Protection Organization. Bulletin OEPP/EPPO Bulletin 40: 401–406.
González, E., Sher, A.A., Tabacchia, E., Masip, A. and Pouline, M. 2015. Restoration of riparian vegetation: A global review of implementation and evaluation approaches in the international, peer- reviewed literature. Journal of Environmental Mana-gement 158: 85–94.
Greenway, M., Dale, P. and Chapman, H. 2003. An assessment of mosquito breeding and control in 4 surface flow wetlands in tropical-subtropical Australia. Water Science & Technology 48: 249–256.
Hashimoto, Y. 2010. Impact of the single cutting in summer on species composition, coverage and species richness in Sicyos angulatus community in the alien species-rich river through the urban area in Japan. Landscape Research Japan Online 3: 32-38. (in Japanese)
Hsiao, A.I. and Huang, W.Z. 1989. Effects of flooding on rooting and sprouting of isolated stem segments and on plant growth of Paspalum distichum L. Weed Research 29: 335–344.
KNA. 2015. The Korean Plant Names Index. Korea National Arboretum. http://www.nature.go.kr/kpni/. Accessed 1 March 2015. (in Korean)
Lee, T.B. 2003. Coloured Flora of Korea. Hyang-munsa, Seoul, Korea. (in Korean)
Liendoa, D., Biurruna, I., Camposa, J.A., Herreraa, M., Loidia, J. and García-Mijangos, I. 2015. Invasion patterns in riparian habitats: The role of anthropogenic pressure in temperate streams. Plant Biosystems 149: 289-297.
Manuel, J.S. and Mercado, B.L. 1977. Biology of Paspalum distichum. I. Pattern of growth and asexual reproduction. Philippine Agriculturist 61: 192-198.
Masters, R.A. and Sheley, R.L. 2001. Principles and practices for managing rangeland invasive plants. Journal of Range Management 54: 502-517.
NIE. 2014. Monitoring of Invasive Alien Species Designated by the Wildlife Protection Act (I). National Institute of Ecology, Seocheon, Korea. (in Korean)
NIER. 2012. Invasive Alien Species. National Institute of Environmental Research, Incheon, Korea. (in Korean)
Noda, K. and Obayashi, H. 1971. Ecology and control of knotgrass (Paspalum distichum). Weed Research, Japan 11: 35-39. (in Japanese)
Naiman, R.J. and Décamps, H. 1997. The ecology of interfaces: Riparian Zones. Annual Review of Ecology and Systematics 28: 621-658.
Oksanen, J. 2013. Multivariate Analysis of Ecological Communities in R: Vegan Tutorial. http://cc.oulu.fi/ ~jarioksa/opetus/metodi/vegantutor.pdf. Accessed 1 April 2015.
Park, S.H. 2009. New Illustrations and Photographs of Naturalized Plants of Korea. Ilchokak, Seoul, Korea. (in Korean)
R Development Core Team. 2013. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org. Accessed 1 April 2015.
Richardson, D.M., Holmes, P.M., Esler, K.J., Galatowitsch, S.M., Stromberg, J.C., Kirkman, S.P., Pysek, P. and Hobbs, R.J. 2007. Riparian vegetation: degradation, alien plant invasions, and restoration prospects Diversity and Distributions 13: 126–139.
Shannon, C.E. and Weaver, W. 1949. The Mathe-matical Theory of Communication. University of Illinois Press, Urbana, USA.
Shin, D.H. and Cho, K.-H. 2001. Vegetation structure and distribution of exotic plants with geomorphology and disturbance in the riparian zone of Seunggi Stream, Incheon. Korean Journal of Ecology 24: 273-280. (in Korean)
Stohlgren, T.J., Bull, K.A., Otsuki, Y., Villa, C.A. and Lee, M. 1998. Riparian zones as havens for exotic plant species in the central grasslands. Plant Ecology 138: 113-125.
Stroh, H.G. 2006. Contribution to the ephemeral wetland vegetation along riverbanks and lakeshores of Western Thrace (NE Greece). Tuexenia 2006: 353-388.
Uchida, T., Nomura, R., Asaeda, T. and Rashid, M.H. 2012. Co-existence of Sicyos angulatus and native plant species in the floodplain of Tama River, Japan. International Journal of Biodiversity and Conservation 4: 336-347.
USDA. 2015. Plants Database. Natural Resources Conservation Service, United States Department of Agriculture. http://plants.usda.gov/. Assessed 5 July 2015.
Van Lill, W.S., Kruger, F.J. and van Wyk, D.B. 1980. The effect of afforestation with Eucalyptus grandis Hill ex Maiden and Pinus patula Schlecht & Cham on stream flow from experimental catch-ments at Mokobulaan, Transvaal. Journal of Hydrology 48: 107–118.
Yang, Y.H., Song, C.K., Park, S.H. and Kim, M.H. 2002. A study on the distribution of naturalized plants of genus Paspalum L. Journal of Subtropical Agriculture and Biotechnology 18: 37-41. (in Korean)
Zavaleta, E., Hobbs, R.J. and Mooney, H.A. 2001. Viewing invasive species removal in a whole- ecosystem context. Trends in Ecology & Evolution 16: 454–459.


