

1. Introduction
2. Survey and Methods
2.1 Survey Site
2.2 Field methods for fish assemblage sampling
3. Results and Discussion
4. Summary and Conclusions
1. Introduction
There are lots of factors that have adversely impacted and disturbed river ecosystems in Korea, including dam lakes and dikes artificially created or installed, river plantation and straightening, aggregate gathering, various civil works and municipal sewage, etc. These factors degrade food chains and ecological habitats for species of fishes that are among primary consumers in river ecosystems, which, in turn, leads to a reduction in their species diversity and habitats.
Species of freshwater fish inhabiting rivers in Korea include 212 species within 39 families of 17 orders. Of them, 61 and 12 species were reported to fall under Korea endemic species and foreign ones, respectively (Kim et al. 2005). Son and Song (2006) reported that 139 species of freshwater fish (within 37 families of 16 orders) inhabited the Geumgang River Basin, while An et al. (1992) reported 70 species within 18 families.
Pseudopungtungia nigra, Pungtungia herzi and Coreoleuciscus splendidus, which belong to Subfamily Gobioninae within family Cyprinidae of Order Cypriniformes, inhabit rivers in Korea (Kim et al. 2005). P. nigra, one of Korea endemic species, is distributed over the Geumgang, Mangyeonggang and Ungcheoncheon River. For their legal protection and management, they were designated as endangered species (Class 1) by the Ministry of Environment in 1998 since their population and range of habitats are on the steady decrease. P. herzi widely spread over in the rivers in Japan and Korea (Kim et al. 2005), and C. splendidus inhabit almost all rivers, except for the Yeongsan River and some other rivers flowing into the East Sea (Kim et al. 2005). Gathering in crowds, these three species inhabit, mainly, riffle or run whose bed substrates consist of materials more sized than gravels. They feed on aquatic insects and attached algae (Kim et al. 2005). Jeon (1977) reported that they were very similar to one another in terms of shape and behavior.
With regard to P. nigra, there were studies on their ecology, egg development (Kim et al. 1991b), spawning and early growth (Lee et al. 2004), and deposition (Kim et al. 2004), including a study on the hybrids of P. nigra and P. herzi (Kim et al. 1991a). Kim and Kang (1990) undertook a systematic study on the skeletal structure and cephalic part of C. splendidus. There were also studies on the micro-distribution (Choi et al. 1990), ecology (Song and Kwon 1993), mutation (Song and Park 2006), and sex chromosome (Kim et al. 2007) of C. splendidus. Hur and Kim (2009) estimated seasonal habitat suitability index (HSI) for water depth, velocity and bed substrates at a downstream site of Yongdam Dam in the Geum River that was required for P. nigra and C. splendidus, and optimal ecological flowrate using physical habitat simulation system (PHABSIM).
This study attempted to address three fish species (P. nigra, P. herzi and C. splendidus) inhabiting the Geumgang River Basin; water depth, velocity, substrate size and microhabitat characteristics required for their habits, including the status of distribution by stream order, were analyzed. It’s expected that the results can be used as basic data for researches on climate change, water improvement projects, aquatic ecosystem con-servations, etc.
2. Survey and Methods
2.1 Survey Site
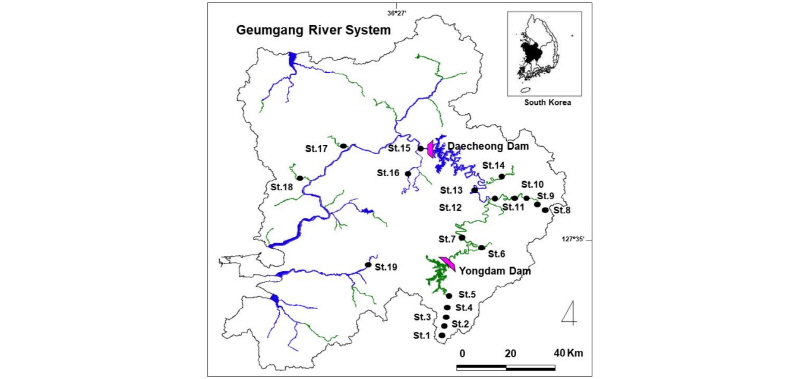
A field survey was carried out midstream and upstream of the Geumgang, Dongjingang and Mang-yeonggang River, the Chogangcheon, Bocheongcheon, Gapcheon, Mihocheon, Musimcheon, Yugucheon, Jicheon, Nonsancheon and Jeongeupcheon stream (Fig. 1). Of those survey sites, however, points where the three species (P. nigra, P. herzi and C. splendidus) didn’t appear were excluded during the result com-pilations; accordingly, results for 19 survey sites were finally compiled.
2.2 Field methods for fish assemblage sampling
Based on the “Guideline to Fish Habitat Monitoring” (Kim and Kim 2009), fish habitat monitoring at the survey sites was carried out. Monitoring processes subsequently followed a preliminary survey, a field survey, and then the compilation and assessment of survey results.
The survey was conducted according to stream order determined in a specific way; the smallest initial tributary of the source of a river is set at “1” in stream order. If this tributary is confluent with another stream with the same order, the confluent stream increases by 1st stream order (Strahler 1952). Each stream order was demarcated using a map with 1 : 75,000 in scale.
The three species of fish were sampled 200 m upstream and downstream of the peak point, equally, for 60 minutes. Fish sampling coverage included pool, riffle and run. A cast net (mesh size: 7 × 7 mm) or kick net (mesh size: 4 × 4 mm) was used depending on river characteristics. When the former was used, it was thrown 15 to 20 times every fish sampling to carry out a quantitative survey. The latter was used for sampling species of fish hiding behind water plants and around cobbles at the right and left river bank. Velocity and water depth at points where those two nets were used, were measured using an electromagnetic flow meter (Model 801, Valeport, UK) and the stake net, re-spectively, and bed substrates are visually inspected with naked eyes. Fish sampling was carried out zigzag from downstream to upstream of the river. Sampled fish were identified in situ, and then released. Fish samples, whose in-situ classification and identification was difficult, were fixed using buffered formalin solution (10%), and then identified in the laboratory. For their classification and identification, studies carried out by Kim et al (2005), Son and Song (2006) and Kim and Park (2007) were referred to.
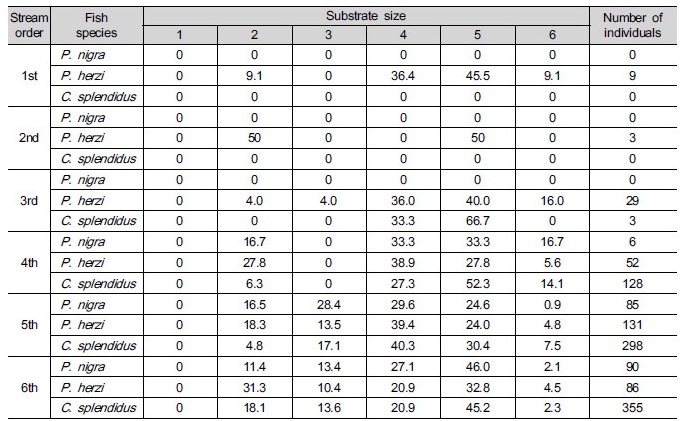
Substrate size were subdivided according to their size: silt (<0.062 mm), sand (0.062 - 2.0 mm), fine gravel (2.0 - 16.0 mm), coarse gravel (16.0 - 64.0 mm), cobbles (64.0 - 256.0 mm), boulders (>256.0 mm) (Woo 2004).
3. Results and Discussion
Three species of fish used in this study belong to family Cyprinidae. It’s reported that they’re similar to one another in terms of shape and behavior, and to Gobiobotia brevibarba, Microphysogobio longidorsalis, Microphysogobio koreensis and Microphysogobio yaluensis in terms of food preferences and habitats (Jeon 1977). Their habitats and distributions, however, have been adversely narrowed due to, for example, changes in river environment and various civil work (Kim et al. 2005).
Tables 1 - 6 shows the appearance and individual distributions of total length, velocity, depth, substrate size and habitat required for P. nigra, P. herzi and C. splendidus according to stream order.
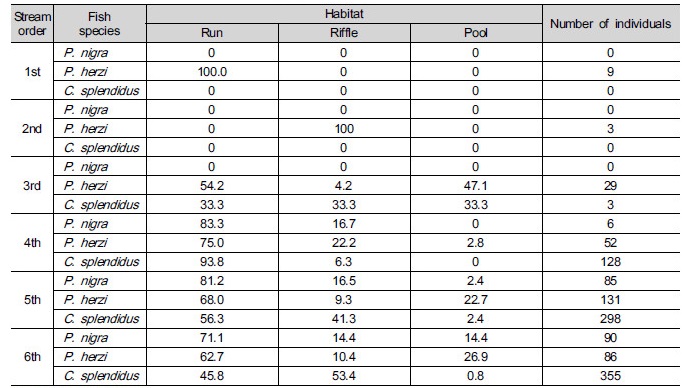
At 9 survey sites (of 19 in total) (47.3%), P. nigra with 181 in number (n=181) appeared, and P. herzi (n=310) appeared at 19 survey sites (100%). C. splendidus with 784 in number appeared at 15 survey sites (78.9%). This suggests that P. nigra, one of en-dangered species, are small and scarce in terms of population and appearance, respectively, when compared with the other two species. P. herzi appeared in all streams, irrespective of their stream order; however, appearance in streams with the 2nd order or lower was found to be scarcer than in the ones with the 3rd order or higher. On the other hand, P. nigra and C. splendidus didn’t appear in streams with the 2nd order or lower. P. herzi were the highest in terms of ap-pearance rate. C. splendidus were also relatively high in appearance rate, but they couldn’t be sampled at points with no riffle. This indicates that their presences are significantly related with the presence of riffle.
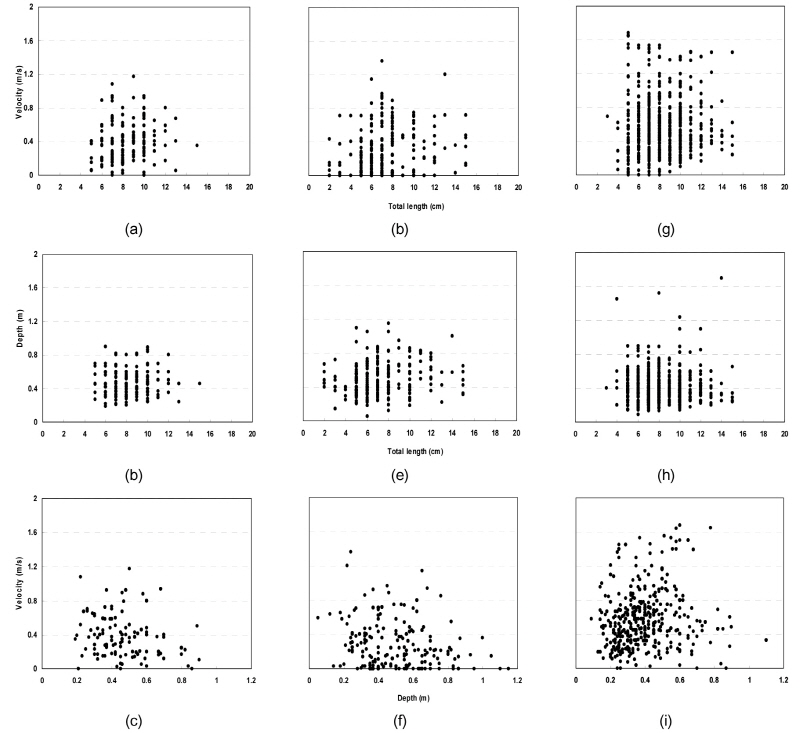
Fig. 2 shows the distributions of velocity and water depth required for P. nigra, P. herzi and C. splendidus according to their length. As seen in the figure, P. nigra with 6 to 11 cm in length were mostly sampled at 0.8 m s-1 or lower in velocity. P. herzi with 5 to 10 cm in length and C. splendidus with 5 to 12 cm in length were mostly sampled at 0.8 m s-1 or lower and 1.5 m s-1 or lower, respectively. P. nigra with 5 to 11 cm in length, and P. herzi and C. splendidus with 5 to 13 cm in length inhabited streams, mainly, with 0.8 m or lower and 0.9 m or lower in water depth, re-spectively. C. splendidus were found to be most widely distributed in terms of the range of water depth and velocity, with P. nigra most narrowly distributed; most of the three species were distributed in streams with 0.8 m or lower in water depth and 0.8 m s-1 or lower in velocity. It was also revealed that C. splendidus were most widely distributed in terms of the range of habitats.
Table 1. Appearance of Pseudopungtungia nigra, Pungtungia herzi, and Coreoleuciscus splendidus at the survey sites of the Geumgang River Basin (The circle means the appearance of each species)
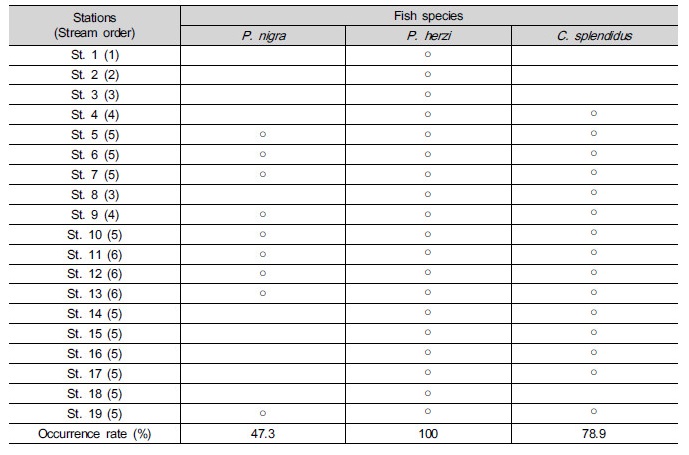 |
Table 2. Frequency (%) of the class of total length of Pseudopungtungia nigra, Pungtungia herzi and Coreoleuciscus splendidus according to the stream order in the Geumgang River Basin
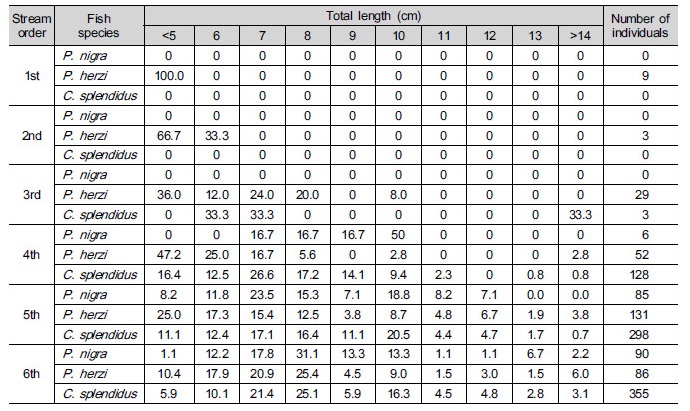 |
Table 3. Frequency (%) of the class of velocity of Pseudopungtungia nigra, Pungtungia herzi and Coreoleuciscus splendidus according to the stream order in the Geumgang River Basin
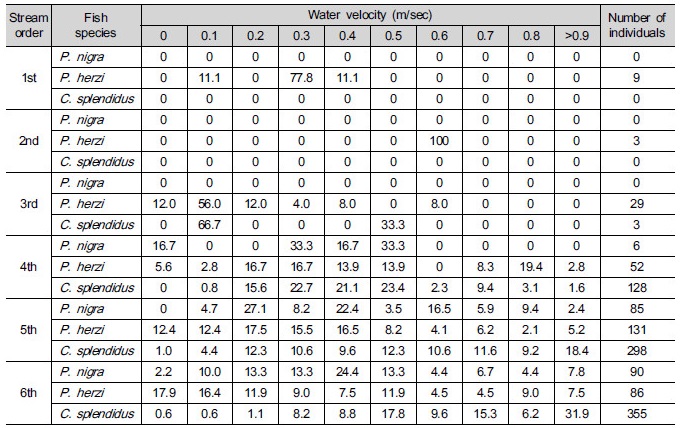 |
Table 4. Frequency (%) of the class of depth of Pseudopungtungia nigra, Pungtungia herzi and Coreoleuciscus splendidus according to the stream order in the Geumgang River Basin
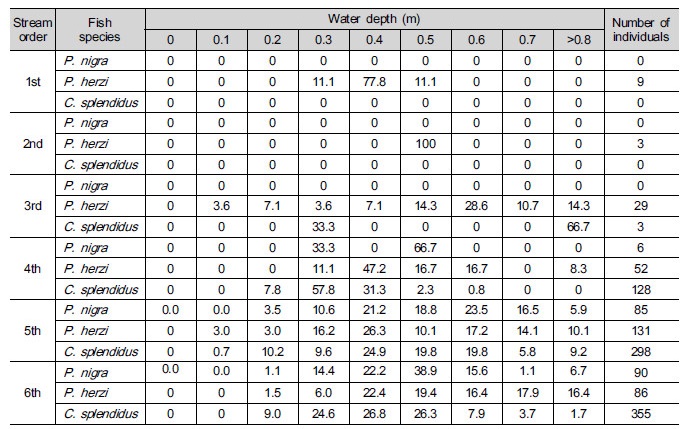 |
Jeon (1977) reported that P. nigra in the GRB inhabited the lower layer (with 0.3 to 1.5 m in water depth) rather than the surface layer. According to his study, species of fish inhabiting the surface layer include Zacco platypus and Z. koreanus, and species of fish inhabiting the lower layer include P. nigra, C. splendidus, Hemibarbus lendirostris and P. herzi. He also found that Coreoperca herzi, Rhinogobius brunneus and Pseudogobio esocinus inhabited the river bed or lived below and around cobbles. Kim et al. (2005) reported that in Mujunamdae and Jujacheon stream, P. nigra with 3 to 4 cm and 6 to 8 cm in length inhabited streams with 0.5 to 1.5 m and 1.5 to 2.5 m in water depth, respectively; his study had showed that they inhabited more deep streams than this study did. It’s judged that this resulted from various differences in, for example, river shape or velocity at sampling points, between the two studies.
t was known that C. splendidus lived, mainly, around riffle with 200 to 300 mm in size of substrates (e.g. cobbles), 0.15 to 1.32 m/s in velocity, and 0.13 to 0.31 m in water depth, and that their size was positively correlated with velocity and water depth (Song and Kwon 1993). Choi et al. (1990) reported that the largest number of species living together with C. splendidus were Z. platypus, followed by P. herzi. They judged that this was because, in Korea, Z. platypus was most widely distributed and highly environment-resistant, not because the two species (i.e. C. splendidus and Z. platypus) had the same habit behaviors. Pielou (1975) termed as “complete association” a case where species “B” necessarily appeared at a point where species “A” lived, but where the latter might not live where the former appeared. In this case, he held that species B was more universal than species A. On the other hand, a term “absolute association” refers to a case where the absence of species A suggests that of species B, and vice versa. With regard to the three species in this study, they were similar in terms of co-existence at points they appeared, although their appearance patterns varied depending on the order of streams they inhabited.
Ludwing and Reynolds (1988) held that an inter- species affinity for coexistence was correlated with habitats, biological and environmental factors, inter- species attraction and repulsion, etc. Jeon (1997) carried out an ecological study on P. nigra and C. splendidus in the GRB, with a focus on their food patterns in riffle and pool. As a result, it was found that C. splendidus preferred aquatic insects, living in riffle. On the hand, P. nigra preferred micro-algae to aquatic insects. They appeared very frequently at points C. splendidus tended to appear, inhabiting pool rather than riffle, and, thereby, avoiding competitive relations with C. splendidus (Jeon 1997). With regard to the habitat patterns of C. splendidus, this study showed findings similar to those of Jeon (1997) study. But P. nigra were likely to frequently appear in runs and the bordering zone of one riffle on another rather than in pool, as shown Table 6 and Fig. 3 (e). It’s judged that this difference from the results of this study is correlated with the fact that the survey period of Jeon (1997) was different from that of this study, and consequently, that there were a significant change in river shape.
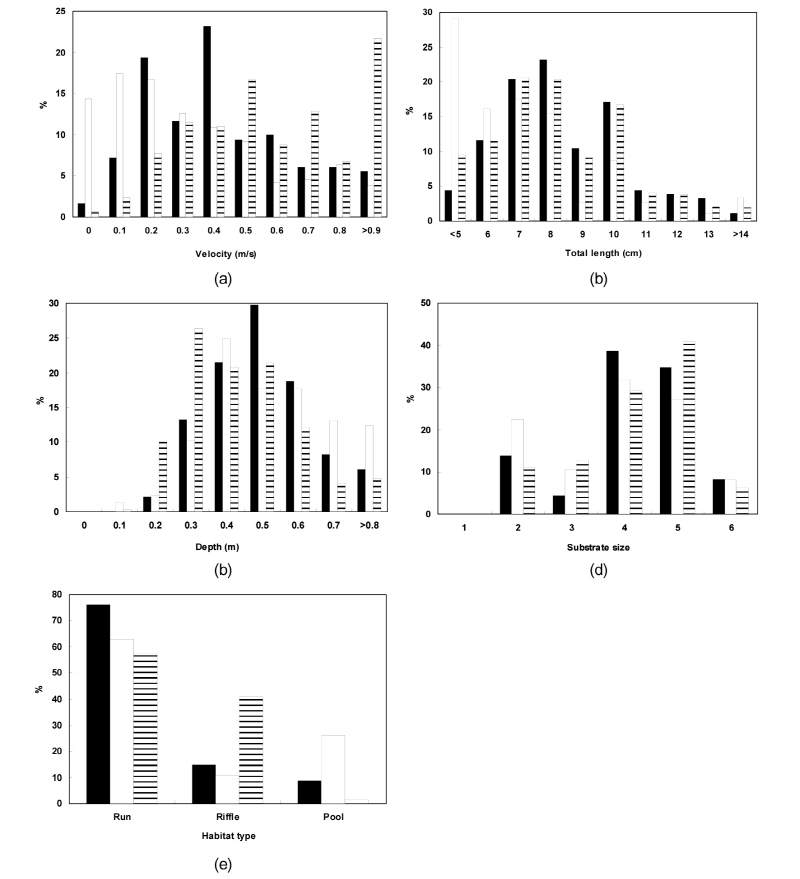
Fig. 3.
Variations of percent to total length, velocity, depth, substrate size and habitat of Pseudopungtungia nigra, Pungtungia herzi and Coreoleuciscus splendidus. Substrate size: 1 (silt, <0.062 mm), 2 (sand, 0.062~2.0 mm), 3 (fine gravel, 2.0~16.0 mm), 4 (coarse gravel, 16.0~64.0 mm), 5 (cobbles, 64.0~256.0 mm), 6 (boulders, >256.0 mm).
Fig. 3 shows how correlated velocity, water depth, substrates and types of habitats required for sampled species of fish, including their length. When it comes to P. nigra, the ones with 7 to 8 cm in length most frequently appeared. They inhabited streams, mainly, with 0.2 to 0.4 m s-1 and 0.5 m in velocity and water depth, respectively. The three species all preferred runs as types of habitats with lots of coarse gravels- cobbles. When it comes to P. herzi, the ones with 5 cm or shorter in length most frequently appeared. They inhabited streams, mainly, with 0 to 0.3 m s-1 and 0.4 m in velocity and water depth, respectively. Finally, C. splendidus with 7 to 8 cm in length occupied the majority of them. They appeared in streams, mainly, with 0.9 m s-1 or higher and 0.3 m in velocity and water depth. Of the three species, C. splendidus lived in streams with the highest velocity. This result is consistent with that of Jeon (1997) and Song and Kwon (1993) study reporting that they inhabited, mainly, riffle. The velocity of riffle at survey points was found to be very high. But they were lower in water depth than run. This indicates that lots of C. splendidus live around riffle.
As seen in the above findings, the three species all inhabited, mainly, the bordering zone of one riffle on another rather than non-stream pool, as reported in Kim et al. (2005) study. It was also revealed that P. nigra and C. splendidus were different in food patterns, and thereby, avoided competitive relations with each other in pursuit of co-existence in microhabitants. These findings are consistent with the results of Jeon (1997). Kim et al. (1991a) reported that there appeared hybrids between P. nigra and P. herzi in a state of nature. This suggests that the three species co-exist in similar habitats. Inhabiting rivers in Korea, they’re known to appear in habitats with good water envir-onment. It was also reported that the characteristics of rivers in which riffle and pool are repeatedly followed by another ones were favorable for their inhabitation and spawning (Kim et al. 2004). As seen in the results of this study, they inhabit, mainly, riffle and are similar in terms of spawning season and other habit behaviors, although there is a slight difference in microhabitats they live in. It’s expected that these results or findings can be used as reference data for researches on climate change and other various river rehabilitation projects in the future.
4. Summary and Conclusions
This study carried out the assessment of microhabitats by stream order Pseudopungtungia nigra, Pungtungia herzi and Coreoleuciscus splendidus in the Geumgang River Basin.
1)Sampled fish totaled up to 1,275. Of them, P. herzi appeared in all streams, irrespective of their stream order, while P. nigra and C. splendidus didn't appear in streams with the 2nd stream order or lower.
2)Most of the three fish species were distributed in rivers with 0.8 m or lower in water depth and 0.8 m s-1 or lower in velocity.
3)Of the three fish species, C. splendidus were most widely distributed in terms of the range of habitats, as is the case with the distribution of their size in terms of water depth and velocity.
4)The three fish species all inhabited, mainly, the bordering zone of one riffle on another rather than non-stream pools, although there was a slight dif-ference in microhabitats they lived in.
5)It’s expected that these results or findings can be used as reference data for researches on climate change and other various river rehabilitation projects in the future.
