

1. 서 론
2. 연구 방법
2.1 조사지 개황
2.2 조사 및 분석 방법
3. 결 과
3.1 식물군락의 다양성과 분포
3.2 생물다양성 현황
3.3 생물상 변화 양상
4. 고 찰
4.1 식생 분포에 따른 습지 생태계 특성
4.2 생물종 분포 특성
5. 결 론
1. 서 론
하구습지는 해양, 하천, 육상 생태계가 맞닿는 전이지대로서, 조수, 염분 구배, 수위 변화 등 복합적인 물리·화학적 요인이 작용하는 동적 생태계이다(Hughes et al. 1998, Weilhoefer 2011, Day et al. 2012). 이와 같은 환경 요인은 조간대의 펄하상, 하중도, 갈대군락 등 다양한 지형과 식생대를 형성하며, 어류, 저서성대형무척추동물, 조류 등 생물종에게 번식지 및 휴식지로서의 핵심 서식처를 제공한다(Dou and Cui 2014, Casazza et al. 2021, Elphick et al. 2022). 이러한 구조적 복합성을 지닌 하구습지는 조수에 적응한 생물의 서식, 염분 구배에 따른 식생대 분포, 해양성·기수성·담수성 생물의 혼재 등 독특한 군집구조를 보이며, 결과적으로 생물다양성의 높은 이질성과 계절적 생산성으로 이어진다(Chi et al. 2018, Zhang et al. 2021). 나아가 하구습지는 단순한 생물 서식처로서의 기능을 넘어, 수질 정화, 영양염류 흡수, 침식 및 퇴적 조절, 홍수 완화 등의 생태계서비스를 제공함으로써 연안 생태계의 기후 회복력 향상에도 중요한 역할을 수행한다(Li et al. 2018, Davidson et al. 2019).
그러나 최근 수십 년간 인구 증가, 도시화, 농지 확장, 기반시설 개발 등의 인위적 교란은 하구 생태계의 심각한 구조적·기능적 변화를 초래하고 있다(Bilkovic et al. 2016). 특히 방파제, 호안 블록, 제방 등 물리적 정비 시설은 침식 및 홍수 피해를 저감하는 기능을 수행하지만, 하천의 곡류 하도 형태를 직선화하고 인접 범람원과의 수문학적 흐름을 차단함으로써 퇴적물 축적 양상과 서식처 공간의 연속성에 중대한 변화를 야기한다(Charlier et al. 2005, Peterson and Lowe 2009). 이러한 구조적 변화는 서식처 단절 및 생물 이동성 저하로 이어지며, 외래종의 확산 등 추가적인 교란 요인을 복합적으로 유발하여 하구습지의 생태적 안정성을 근본적으로 위협한다(Boys et al. 2012, Laurance et al. 2012). 즉 하구습지의 수문학적 연결성(hydrological connectivity)은 물질·에너지·생물의 이동을 매개하며, 생물다양성 유지에 핵심적인 역할을 수행하므로(Taylor et al. 1993, Erős et al. 2012), 하구습지 보전 전략은 단순한 보호구역 지정 차원을 넘어 단절된 수문·생태 연결성의 회복과 지속적 유지를 중점적으로 고려해야 한다.
경상남도 하동군에 위치한 주교천은 섬진강으로 유입되는 지방하천으로, 이 중 본 연구의 대상지인 주교천하구습지는 섬진강 합류 지점인 말단부에 위치하며 남해의 광양만으로 흘러들어가는 열린 하구로서의 특징을 가진다. 이 지역은 해수와 담수가 만나는 조수 간만대로, 퇴적지형, 갈대군락, 염습지 등이 공존하는 전형적인 전이지대를 형성하고 있다(Lee et al. 2025). 인근 광양만 일대는 1970년대 이후 광양제철소 건설과 산업단지 조성에 따라 대규모 간척 및 매립사업이 진행되면서 자연 해안선의 대부분이 인공화되었고(Roh and Lee 2006), 주교천하구습지는 이러한 개발 압력의 경계에 놓인 상대적으로 보전된 습지 공간으로 기능하고 있다. 주교천은 주기적인 하천정비 기본계획에 따라 제방 구축과 하도 정비가 이루어졌고, 이 과정에서 자연곡류 구간의 직강화와 수문 흐름의 변화가 있었던 것으로 판단된다(Gyeongsangnam-do 2006). 그러나 이 지역에 대한 생물학적 모니터링 및 생태학적 영향 평가는 거의 전무하며, 인접한 섬진강 하구에서 어류, 저서성대형무척추동물, 식생 및 플랑크톤 등을 대상으로 다수의 조사가 이루어진 것과는 대비된다. 정량적 생태자료의 부재는 주교천하구습지의 국지적 특성과 생태적 연속성을 규명하는데 어려움을 초래하고 있으며, 이에 대한 정밀 생태 진단과 보전 가치 평가가 요구된다.
이에 본 연구는 국립생태원에서 수행한 “하구 생태계 조사” 결과를 바탕으로 주교천하구습지의 생태계 현황 및 특성을 종합적으로 파악하고, 습지보호지역 지정 가능성을 검토하는 것을 목적으로 하였다. 이를 위해 (1) 염도 구배 및 지형 특성에 따라 구분된 권역별 식물군락과 멸종위기 야생생물, 생태계교란 생물을 포함한 생물다양성 현황을 파악하고, (2) 권역별 식생 및 지형 특성에 따른 생물상 분포 현황을 분석하여, (3) 주교천하구습지의 생태적 보전 가치를 평가하고 이를 바탕으로 보전 및 관리 방안을 제안하고자 한다.
2. 연구 방법
2.1 조사지 개황
본 연구의 대상지인 주교천하구습지는 경상남도 하동군 고전면, 금남면, 금성면을 관류한 후 섬진강과 합류하여 남해 광양만으로 유입되는 지방하천인 주교천의 하류부에 위치한다. 하구습지가 지닌 생태적 특성을 온전하게 평가하기 위해, 기수역이 형성되는 구간을 중심으로 조사 범위를 설정하였다. 종적 범위는 섬진강하구습지가 포함된 하류부에서부터 만조 시 해수 유입이 제한되는 고전천 합류 지점(고전면 대덕리 인근)까지이며, 횡적 범위는 제내지 일부 경작지를 포함하였다. 특히 섬진강하구습지는 국가하천이자 광양만으로 유입되는 주요 하구로서 주교천과의 생태적 연계성을 고려하여 조사 범위에 포함하였다. 전체 조사 대상지의 총 연장과 면적은 각각 4.2 km와 1,764,519 m2로, 이는 주교천 유역면적(81.3 km2)의 약 2.2%에 해당한다(Fig. 1).
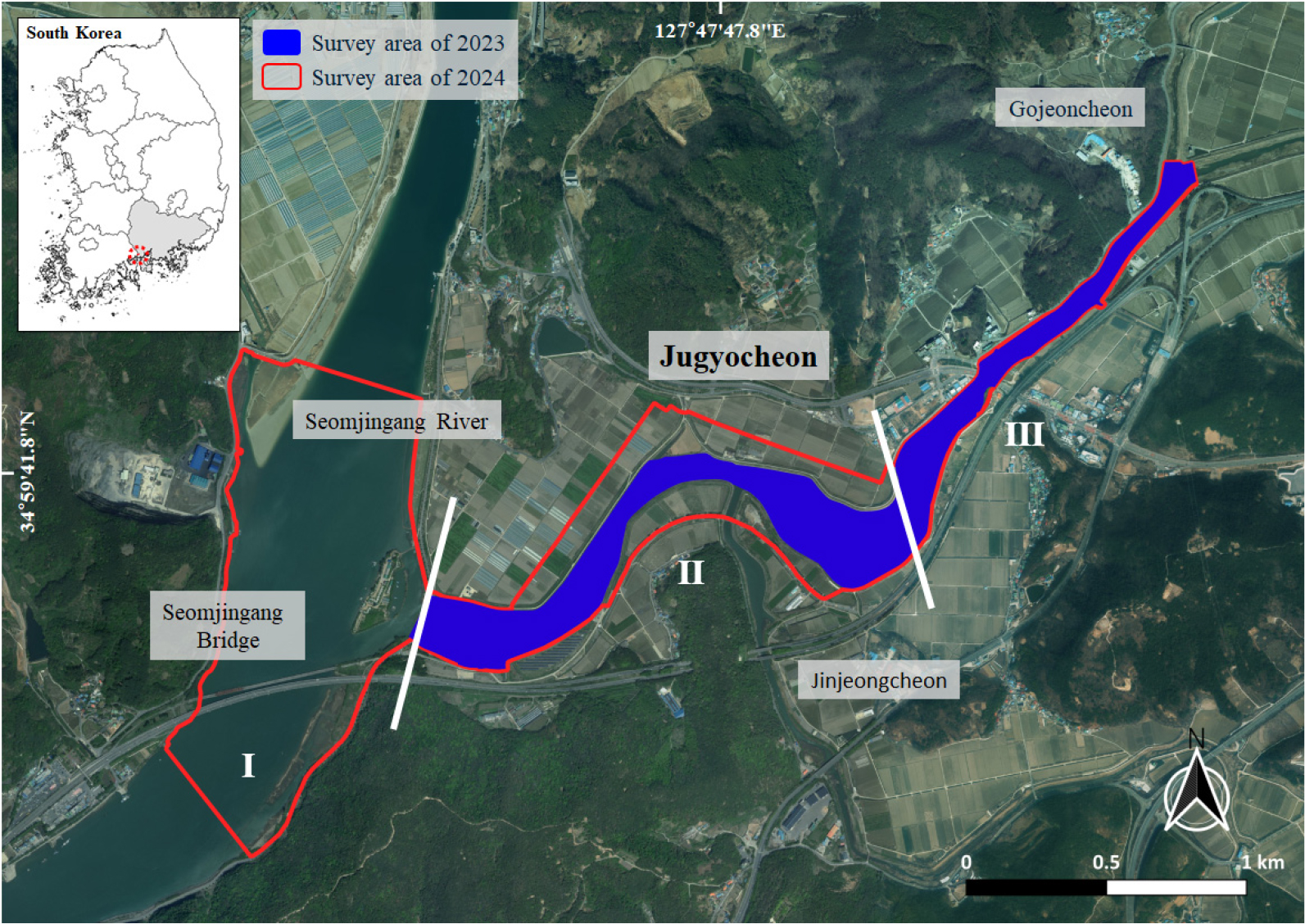
Fig. 1.
Map showing the study site of the Jugyocheon Estuarine Wetland, Korea. Red lines on the map indicate survey areas for 2024, while blue areas on the map indicate survey areas from 2023. White lines delineate reaches Ⅰ (Lower reach), Ⅱ (Middle reach), and Ⅲ (Upper reach) of the 2024 survey area. Black lines on the keymap indicate borders of provinces. Red dashed line marks the location of the Jugyocheon Estuarine Wetland.
지형적 조건과 염도 구배 등을 고려하여 조사 대상지는 상류, 중류, 하류의 3개 소권역으로 구분하였으며, 각 권역별로 분야별 현장조사를 수행하였다. Ⅰ구간은 섬진강 하구의 본류에 해당하며, 해수의 영향을 강하게 받는 기수역으로 조석 변동에 따른 수위 변화가 크다. 염도는 10.7 – 12.5%로 가장 높은 값을 보였으며, 하도 양안에는 점토질 간석지가 잘 발달하였다. 주교천과 섬진강이 합류하는 지점에는 대규모 하중도(조개섬)가 형성되어 있었으나, 현재는 캠핑장으로 이용되고 있다. Ⅱ구간은 곡류 형태의 하천 지형이 비교적 잘 보전되어 있으며, 광범위한 사력퇴적지가 분포하고 있다. 해당 구간에서는 갈대군락이 단일식생으로 고밀도 우점하고 있다. 진정천 합류부를 기준으로 염도가 10.9%에서 0.5%로 급격하게 감소하였으며, 인접 지역은 주로 농경지로 이용되고 있다. Ⅲ구간은 염도가 0.1 – 0.7%로, 해수의 영향을 거의 받지 않는 담수역에 해당한다. 하폭은 약 50 – 60m로 급격히 좁아지며, 유로는 직선형으로 길게 이어지고 있었다. 상류에 설치된 보의 영향으로 유속이 감소하면서 사력퇴와 분류하도와 같은 퇴적지형이 발달하였다.
본류 구간의 퇴적물은 세립질 점토(≤0.002 mm)부터 조립질 거력(>256 mm)에 이르기까지 다양한 입경 범위를 포함하고 있으며, 상류와 하류 간 입경 조성에는 뚜렷한 경향성은 관찰되지 않았다(NIE 2024). 토성은 사토, 사양토, 양토, 미사질양토 등 여러 유형이 혼재하였으며, 전반적으로는 실트 함량이 높은 미사질양토가 우세하였다. 기후 특성 분석을 위해, 주교천하구습지와 인접한 진주 기상관측소에서 1995년부터 2024년까지 관측된 자료를 확인한 결과, 해당 기간의 연평균 기온은 13.5°C, 연평균 강수량은 1,552 mm로 나타났다(KMA 2024).
2.2 조사 및 분석 방법
주교천하구습지의 생물다양성과 생태계 특성을 파악하기 위하여, 2024년 2월부터 11월까지 총 9개 생물 분야(식생, 식물상, 포유류, 조류, 어류, 양서·파충류, 육상곤충, 담수무척추동물, 기수무척추동물)를 대상으로 현장조사를 실시하였다. 조류는 철새 도래지로서의 생태적 가치를 고려하여 겨울철 조사를 포함하였으며, 기타 분야는 생태적 특성과 계절성을 반영하여 조사시기를 차등 적용하였다(Table 1). 현장조사는 각 분야의 전문가에 의해 수행되었으며, 전반적인 방법론은 내륙습지 조사지침(NIE 2020)에 제시된 분야별 조사 기준을 준용하였다. 식생조사는 Braun-Blanquet(1965)의 식물사회학적 방법론에 따라 실시되었으며, 피도는 9단계 통합우점도 체계를 적용하여 기재하였다. 주요 식생 단위의 대표성이 확보할 수 있는 지점을 선정하여 총 31개의 식생조사표를 구축하였고, 실내 분석을 통해 식생 유형을 분류하였다. 이후, 정밀 항공사진 분석과 현장조사 결과를 종합하여 상관수준에서 식물군락명을 부여하였으며, QGIS 프로그램을 활용하여 현존식생도를 작성하고 군락별 분포면적을 산정하였다(QGIS Development Team 2025). 식물상 조사는 도보를 통한 선조사법으로 수행되었으며, 출현종을 직접 관찰하여 목록화하였다. 어류는 조사정점에서 투망과 족대를 활용하여 채집하였고, 양서·파충류는 포충망과 뱀집게를 이용한 직접 확인과 울음소리를 통한 간접 확인을 병행하였다. 육상곤충 조사는 주간에는 쓸어잡기, 채어잡기 방식의 포충망 채집, 야간에는 등화채집(light trap)을 병행하여 수행하였다. 포유류는 배설물, 족적 등 흔적을 확인하는 직접 조사와 함께 소형종과 중·대형종을 구분하여 무인센서카메라를 설치·운영함으로써 지속적인 모니터링을 수행하였다. 조류는 쌍안경과 망원경 등을 이용하여 정점 및 선을 따라 관찰하였으며, 담수 및 기수무척추동물은 D-frame net, hand net, 삽 등을 사용하여 정점에서 정량 및 정성 채집을 병행하였다. 이 중 식물, 양서·파충류, 육상곤충, 포유류, 조류는 전 구간에 걸쳐 전수조사를, 어류와 무척추동물은 대표 정점을 선정하여 조사하였다.
Table 1.
Survey period according to survey order for taxonomic group
한편, 습지의 생태적 건강성 평가는 식생, 어류, 저서성대형무척추동물을 대상으로 실시하였으며, NIER (2024)에서 제시한 평가지수를 활용하여 A – E 등급의 5단계로 구분하였다. 하구습지의 온전한 식생분포를 파악하기 위해 하천 본류 구간인 제외지에 분포하는 식물군락만을 대상으로 범위를 설정하였으며, 하구수변식생평가지수(Korea Estuary Riparian Vegetation Index; KERVI)를 적용하여 식생환경의 온전성, 습지식물군락의 다양성, 외래식물·육상식물·습지식물군락의 면적비율을 종합적으로 고려하였다. 어류와 저서성대형무척추동물은 각각 하구어류평가지수(Korea Estuary Fish Assessment Index; KEFAI) 및 하구저서성 대형무척추동물평가지수(Korea Estuary Benthic Macroinvertebrates Index; KEBMI)를 적용하여 종다양도, 생태적 기능군의 구성, 출현종수 등을 기준으로 평가하였다. 또한 주교천하구습지에서의 생물상 변화 양상을 파악하기 위해, 2023년에 수행된 하구습지 기초조사 결과를 활용하였다(NIE 2023). 당시 조사는 섬진강 본류를 제외한 수역 중심으로 이루어졌으며, 육역 공간은 조사 대상에 포함되지 않았다(Fig. 1). 이에 따라 양서·파충류, 어류, 조류, 담수무척추동물, 기수무척추동물 등 기초조사에서 수행된 분류군에 한하여 본 조사 결과와 비교 분석을 진행하였다. 다만, 조사 강도와 시기적 차이에 따른 환경 요인에 대한 보완 자료가 부족하여 생물상 변화 해석에는 일정 수준의 불확실성이 수반될 수 있다. 마지막으로 식생 및 생물 분포 특성을 종합적으로 분석하여 습지의 생태적 가치를 평가하고 보호지역으로서의 지정 가능성과 이에 따른 보전 방안을 제안하였다.
3. 결 과
3.1 식물군락의 다양성과 분포
주교천하구습지에는 하천의 지형적 구배, 수문학적 연결성, 그리고 인위적 간섭 등의 복합적인 환경 요인에 따라 총 5개 상관식생형의 18개 식물군락이 분포하는 것으로 확인되었다. 조사 구역은 인공제방을 경계로 식물군락의 유형과 토지이용 특성이 뚜렷하게 구분되었다. 제방 외측(제외지)에는 정수 및 유수역 환경에 적응한 다년생 초본 식생 등 자연성이 높은 군락이 우점하였으며, 내측인 제내지에는 경작과 조경식재 등 인위적 간섭의 영향으로 경작지식생, 대상식생, 기타식생이 주로 분포하였다(Table 2). 식생의 수분 요구도를 기준으로 식물군락을 분류한 결과, 절대습지식물(obligate wetland plant; OBW) 우점군락 4개, 임의습지식물(facultative wetland plant; FACW) 우점군락 1개, 양생식물(facultative plant; FAC) 우점군락 5개, 절대육상식물(obligate upland plant; OBU) 우점군락 7개로 나타났으며, 경작 목적의 식물군락인 벼군락(Oryza sativa community)은 분석에서 제외하였다.
Table 2.
Vegetation diversity in the Jugyocheon Estuarine Wetland, Korea
하구습지 전구간에 걸쳐 정수역에 발달하는 다년생 초본식생인 갈대군락(Phragmites australis community)이 총 291,425 m2에 이르는 넓은 면적으로 분포하고 있으며, 단일 종에 의한 우점 양상이 뚜렷하게 확인되었다(Fig. 2). 특히 수심 10 cm 이상의 수변부에서 주로 분포하며, 중류 지역의 사력퇴가 발달한 지점을 중심으로 약 100,000 m2 이상 대규모의 단일군락을 형성하고 있다. 천일사초군락(Carex scabrifolia community)은 갈대군락과 인접한 유수역에 분포하고 있으며, 유속에 의한 물리적 교란이 상대적으로 적은 안정적인 서식처 환경에서 유지되는 것으로 보인다. 일반적으로 하구습지는 염도, 조석 등 환경 요인의 영향으로 식생 다양성이 낮게 나타나는 것이 일반적이나, 주교천 본류 구간은 이보다 더 단순화된 식생 구조를 보이고 있었다.
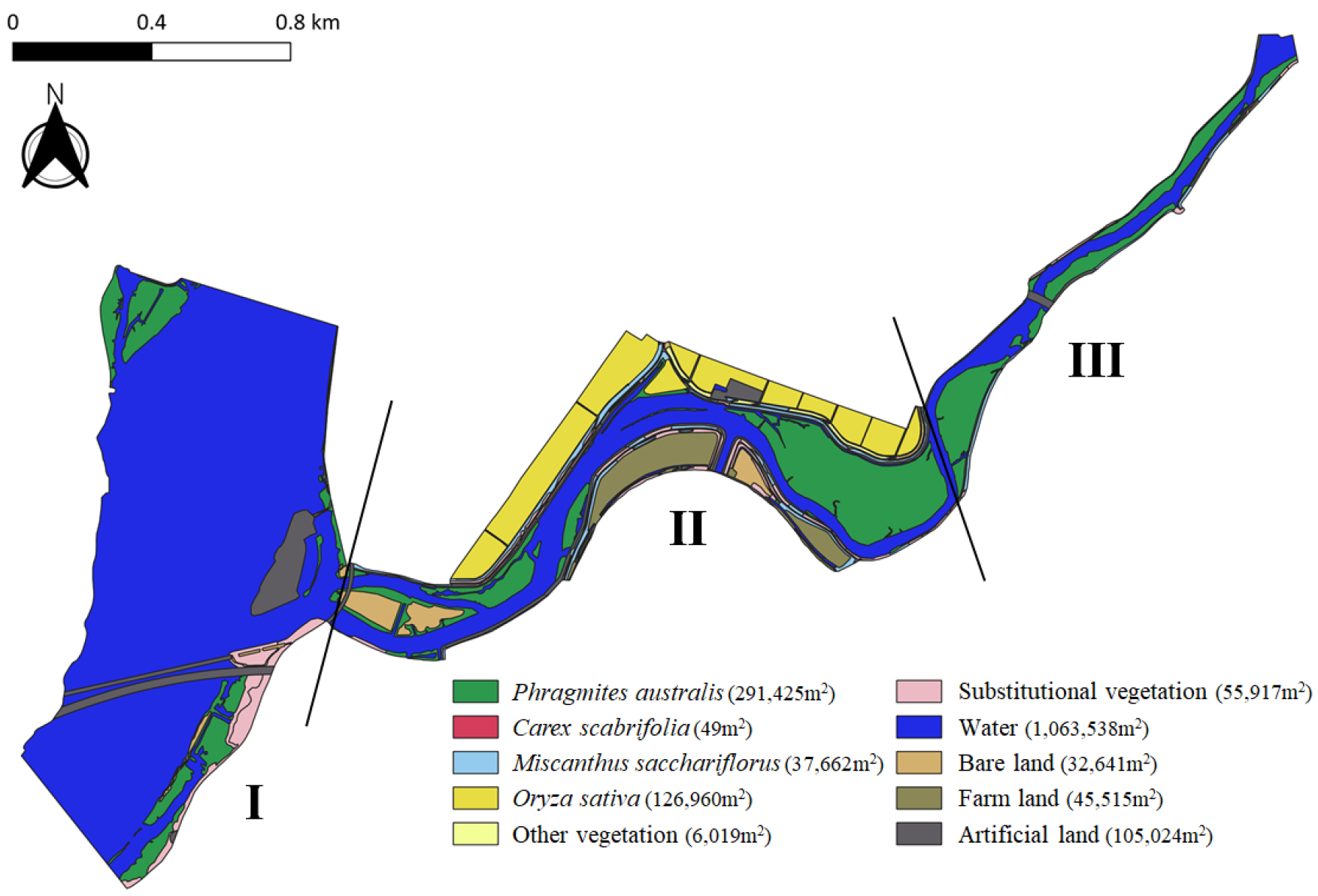
제방 내·외측 사면에서는 물억새군락(Miscanthus sacchariflorus community)이 띠 모양으로 길게 대상분포하고 있었다. 일반적으로는 장마 후 환삼덩굴군락(Humulus japonicus community)이 물억새군락을 피복하여 생육을 억제하는 양상이 나타나나, 주교천에서는 늦여름까지도 물억새군락이 지속적으로 유지되고 있는 것이 특징이다. 이 외에도 하천 범람을 방지하기 위해 조성된 급경사의 제방 내측 사면에서는 칡군락(Pueraria lobata community), 환삼덩굴군락, 큰김의털-털빕새귀리군락(Bromus tectorum-Festuca arundinacea community) 등의 초본성 식물군락과 더불어, 족제비싸리군락(Amorpha fruticosa community), 아까시나무군락(Robinia pseudoacacia community) 등 인위적으로 식재된 관목성 군락이 패치 형태로 산발적으로 분포하고 있었다. 이러한 양상은 제내지 내에서 인위적 교란에 의해 자연적인 군락 이외의 식생이 형성되면서, 일정 부분 식생의 이질성이 증가하고 있음을 시사한다.
3.2 생물다양성 현황
주교천하구습지의 생물다양성 현황을 8개 주요 생물 분류군을 대상으로 분석한 결과, 총 1,074종이 확인되었다(Table 3, Table 4). 이 중 멸종위기 야생생물은 포유류, 조류, 저서성대형무척추동물 분류군에서 총 15종이 출현하였다. 포유류에서는 Ⅰ급 수달(Lutra lutra), Ⅱ급 담비(Martes flavigula), 삵(Prionailurus bengalensis) 등 3종이 관찰되었고, 조류에서는 Ⅰ급 노랑부리백로(Egretta eulophotes) Ⅱ급 붉은배새매(Accipiter soloensis), 큰기러기(Anser fabalis), 흰목물떼새(Charadrius placidus), 잿빛개구리매(Circus cyaneus), 큰고니(Cygnus cygnus), 새호리기(Falco subbuteo) 등 7종이, 저서성대형무척추동물 분야에서는 Ⅱ급 흰발농게(Austruca lactea), 갯게(Chasmagnathus convexus), 기수갈고둥(Clithon retropictus), 대추귀고둥(Ellobium chinense), 붉은발말똥게(Sesarmops intermedius) 등 5종이 기록되었다. 구간별 종수는 조석 및 해수 유입의 영향을 받는 하류(Ⅰ구간, 562종) 및 중류(Ⅱ구간, 525종)에서 상대적으로 높았으며, 담수 환경이 우세한 상류(Ⅲ구간)에서는 486종으로 다소 낮게 나타났다. 분류군별 출현 양상은 서식처 특성과 염도 조건에 따라 상이하였다. 양서·파충류와 담수무척추동물은 염도 영향이 적은 Ⅲ구간에서 종풍부도가 높았던 반면, 육상곤충, 조류, 기수무척추동물은 기수역이 발달한 Ⅰ구간에서 높은 종풍부도를 보였다. 그 외의 분류군은 구간별 차이가 뚜렷하지 않았으며, 각 분야별 상세 결과는 다음과 같다.
Table 3.
Changes in the number of taxa in the upper, middle, and lower reaches of the Jugyocheon Estuary Wetland in Korea from 2023 to 2024
| Taxonomic group | 2024 | 2023** | |||
| Ⅰ(Lower) | Ⅱ(Middle) | Ⅲ(Upper) | Total | ||
| Plant* | - | - | - | 194 | - |
| Amphibian/reptile | 0/0 | 5/4 | 6/4 | 6/5 | 2/0 |
| Fish | 16 | 12 | 15 | 27 | 19 |
| Bird | 43 | 46 | 25 | 67 | 25 |
| Mammal | 10 | 8 | 10 | 14 | - |
| Benthic invertebrate (fresh water) | 11 | 19 | 22 | 30 | 7 |
| Benthic invertebrate (sea water) | 29 | 19 | 14 | 33 | 16 |
| Terrestrial insect | 453 | 412 | 390 | 698 | - |
| Total | 562 | 525 | 486 | 1,074 | 69 |
Table 4.
The list of endangered species designated by the Korea Ministry of Environment in the Jugyocheon Estuarine Wetland, Korea
관속식물은 56과 141속 179종 1아종 13변종, 1품종 등 총 194분류군이 확인되었다. 간조 시 노출되는 Ⅰ·Ⅱ구간의 염습지에서는 나문재(Suaeda glauca), 가는갯는쟁이(Atriplex gmelinii), 칠면초(Suaeda japonica) 등 9분류군의 염생식물이 분포하였다. 또한 말즘(Potamogeton crispus), 가래(Potamogeton distinctus), 개구리밥(Spirodela polyrhiza), 모새달(Phacelurus latifolius) 등 다양한 생육형의 습지식물 32분류군이 확인되었다. Ⅱ구간은 곡류 하천지형과 대규모 사력퇴가 발달하며, 이에 따라 갈대가 우점하고 있으며, 제내지에 위치한 소규모 수변습지에서는 부들(Typha orientalis), 질경이택사(Alisma orientale) 등도 관찰되었다. Ⅲ구간은 인공제방, 수중보 영향으로 식물 정착이 제한적이나, 유량이 안정화되는 고남교 인근의 사력퇴를 중심으로 갈대, 줄(Zizania latifolia), 달뿌리풀(Phragmites japonica) 등의 정수식물이 일부 정착하고 있었다.
양서·파충류는 양서류 3과 6종, 파충류 3과 5종으로 총 6과 11종 210개체가 확인되었다. 우점종과 아우점종은 각각 생태계교란 생물인 황소개구리(Rana catesbeiana)와 붉은귀거북(Trachemys scripta)으로 나타났다. 출현 개체는 주로 제내지에 위치한 농경지와 둠벙, 인접 산림 등 다양한 서식환경이 혼재된 지역에서 관찰되었으며, 하천 본류의 제외지 구간은 해수 유입에 따른 기수역이 형성되어 양서·파충류의 서식이 매우 제한적이었다. 특히, 조수의 영향을 직접적으로 받는 하류부(Ⅰ구간)에서는 출현이 전혀 확인되지 않았다.
어류는 총 10과 27종 1,512개체가 확인되었으며, 우점종은 송사리(Oryzias latipes, 297개체), 아우점종은 민물검정망둑(Tridentiger brevispinis, 246개체)으로 나타났다. 펄질 하상이 광범위하게 발달하고 해수 영향이 지속되는 환경으로 인해, 짱뚱어(Boleophthalmus pectinirostris), 큰볏말뚝망둥어(Periophthalmus magnuspinnatus), 점줄망둑(Acentrogobius pellidebilis) 등 다양한 망둥어과 어류를 포함하여 해산·기수·회유성 기능군의 어류 19종이 확인되었다. Ⅰ구간에서는 해산성 어종의 치어가, Ⅲ구간에서는 1차담수어 및 회유성 어종이 주로 관찰되었다. 또한 2021년 남해군에서 국내 신종으로 보고된 흰점실고기(Hippichthys penicillus)가 섬진강 수계에서는 최초로 확인되었으며, 블루길과 배스 등 생태계교란 생물은 출현하지 않았다.
육상곤충은 총 13목 121과 698종이 확인되었으며, 구간별로는 Ⅰ구간 453종, Ⅱ구간 412종, Ⅲ구간 390종이었다. 나비목이 436종(62.5%)으로 가장 높은 비율을 차지하였고, 이어 딱정벌레목(12.0%), 노린재목(10.7%) , 메뚜기목(4.6%) 순이었다. 기후변화 지표종은 각시메뚜기(Patanga japonica), 말매미(Cryptotympana atrata), 푸른아시아실잠자리(Ischnura senegalensis) 등 6종, 생태계교란 생물은 등검은말벌(Vespa velutina nigrithorax), 미국선녀벌레(Metcalfa pruinosa), 갈색날개매미충(Ricania sublimata) 3종이 출현하였다. 특히 Ⅰ구간은 산림, 섬진강과의 합류 등 인접한 자연환경과의 연결성이 높아 상대적으로 높은 종풍부도를 보였으며, 2024년에 미기록종으로 보고된 검은눈그물나비(Neope muirheadii)의 유충과 성충이 모두 확인되었다.
조류는 총 30과 67종 1,557개체가 조사되었으며, 우점종은 제비(Hirundo rustica, 252개체), 아우점종은 흰뺨검둥오리(Anas poecilorhyncha, 215개체)였다. 도요·물떼새, 오리류, 백로류 등 수조류는 31종 836개체로, 전체 개체수의 절반 이상을 차지하였다. Ⅰ ~ Ⅱ구간에서는 갈대군락, 논습지 등 다양한 서식처와 철새도래지(광양만, 갈사만)와의 연계성으로 인해 약 45종의 높은 종풍부도가 확인되었으며, Ⅲ구간은 좁은 수로형 하천 형태의 단순한 경관 특성으로 출현종수가 25종으로 낮았다. 노랑부리백로, 큰기러기, 흰목물떼새 등 멸종위기 야생생물 7종이 확인되었으며, 하상 사력퇴, 주변 논습지와 수목대 등 각각 선호하는 서식 환경에서 휴식 및 섭식 활동이 관찰되었다.
포유류는 총 5목 10과 14종이 조사되었으며, 흔적조사 및 카메라 트랩을 통해 등줄쥐(Apodemus agrarius, 245회), 고라니(Hydropotes inermis, 131회), 수달 (105회)이 높은 빈도로 관찰되었다. 밀생한 갈대군락이 분포한 Ⅱ구간에서는 등줄쥐, 작은땃쥐(Crocidura shantungensis), 멧밭쥐(Micromys minutus) 등 소형 포유류가 집중적으로 출현하였으며, 수달은 중·하류 구간에서 최대 4개체가 동시에 관찰되는 등 번식과 먹이활동을 활발히 수행하는 것으로 확인되었다. 삵은 하구습지 전 구간에서, 담비는 산림과 연결된 이동통로로서 습지를 이용하는 모습이 관찰되었다(Fig. 3).

Fig. 3.
Distribution map of endangered wildlife in the Jugyocheon Estuarine Wetland, Korea. Point symbols indicate observation sites of individual species. Polygons denote potential habitat zones of brackish invertebrate species, inferred from scattered occurrences across multiple survey sections. These zones were mapped as continuous areas for illustrative clarity, though actual distributions were spatially intermittent.
담수무척추동물은 총 23과 30종이 조사되었으며, 이 중 수서곤충류가 40% (12종)를 차지하였다. 우점종은 실지렁이(Limnodrilus gotoi), 아우점종은 줄새우(Palaemon paucidens)였다. Ⅰ구간에서 11종, Ⅱ구간 19종, Ⅲ구간 22종으로 상류로 갈수록 종풍부도가 증가하는 경향을 보였다. Ⅰ구간은 높은 염도와 조석 영향으로 수서곤충류가 전혀 출현하지 않았으며, Ⅱ구간 19종, Ⅲ구간 22종으로 상류로 갈수록 출현 종수가 증가하였다. 이는 해수의 영향이 줄어들어 수서곤충류의 서식 조건이 점진적으로 회복되는 양상이 반영된 것으로 판단된다.
기수무척추동물은 멸종위기 야생생물 5종을 포함해 총 18과 33종이 확인되었다. 절지동물문 연갑강이 17종으로 가장 높은 종다양성을 보였으며, 연체동물문 복종강과 이매패강 순이었다. Ⅰ구간 29종, Ⅱ구간 19종, Ⅲ구간 14종으로 상류로 갈수록 종풍부도가 감소하는 경향을 보였다. 특히 갈대가 넓게 분포한 사력퇴에서 말똥게(Chiromantes dehaani), 붉은발말똥게 등 대규모 군집이 관찰되었으며, 기질 유형에 따라 칠게(Macrophthalmus japonicus), 엽낭게(Scopimera globosa) 등이 선택적으로 출현하였다. 기수갈고둥은 염도(0.1 ~ 0.3 %)가 낮고 잔자갈(2 ~ 16mm)이 분포하는 Ⅲ구간에서, 흰발농게와 대추귀고둥은 조간대 상부의 갈대군락이 형성된 Ⅰ구간 및 Ⅱ ~ Ⅲ 구간에서 각각 대규모로 서식하고 있었다(Fig. 3).
3.3 생물상 변화 양상
2023년에 수행된 주교천하구습지 조사 결과와 비교하여 생물상 변화 양상을 분석하였다(Table 3). 본 비교는 동일 조사 구간 및 분류군(어류, 조류, 양서·파충류, 담수무척추동물, 기수무척추동물)에 한하여 수행되었다. 총 출현 종수는 2023년 69종에서 2024년 136종으로 67종이 증가하였고, 어류를 제외한 대부분의 분류군에서 종수 증가가 뚜렷하게 나타났다. 특히 기수무척추동물을 제외한 분류군에서 2023년에 기록된 종의 70 - 100%가 동일하게 재출현하였으며, 종수 증가는 대부분 2024년에 새롭게 출현한 종에 기인하였다. 다만 2023년 조사는 기초조사 성격으로 2024년 보다 조사 강도 및 빈도가 상대적으로 낮았고 조사 시기의 기상 및 수질 등 환경 요인에 대한 정보도 부족하여 두 시기의 결과를 정량적으로 직접 비교하는 것은 한계가 있었다. 따라서 출현 종수 증가가 반드시 실제 생물다양성의 증가를 의미한다고 단정하기는 어렵다. 그럼에도 불구하고, 본 비교 결과는 주교천하구습지가 다양한 생물군이 출현할 수 있는 서식 여건을 지닌 지역임을 시사하는 기초자료로 활용될 수 있으며, 향후 생물다양성의 실질적인 변화 추이를 파악하기 위해서는 동일 구간·동일 시기·표준화된 조사 설계에 기반한 반복적이고 장기적인 모니터링이 필요할 것이다.
분야별 주요 변화 양상으로 양서·파충류에서는 생태계교란 생물인 황소개구리가 2023년에 이어 지속해서 출현하였으며, 제내지에 위치한 담수 미소서식처가 포함되어 종수 증가에 기여하였다. 어류는 송사리(Oryzias latipes), 큰볏말뚝망둥어(Periophthalmus magnuspinnatus) 등 기수성 어종 포함 8종이 새롭게 확인된 반면, 문절망둑, 왜몰개, 납지리, 잉어 등 일부 기존 출현종은 관찰되지 않았다. 이는 조사 시기 또는 환경 조건의 변화에 따른 일시적인 부재로 판단된다. 조류는 2023년에 확인된 모든 종이 재출현하였으며, 수조류 및 산림성 조류 등 26종이 추가로 확인되었다. 다양한 생태적 지위를 지닌 조류가 폭넓게 출현한 점은, 조사 범위 및 빈도의 증가와 더불어 대상지의 복합적인 서식환경이 반영된 결과로 판단된다. 담수무척추동물은 기존 7종에서 30종으로 크게 증가하였으며, 이는 조사 구간 내 담수 환경의 비중 확대에 따른 결과로 판단된다. 기수무척추동물은 과거 출현 종의 절반 이상이 교체되어 종 조성 변화가 가장 두드러졌으며, 특히 멸종위기 야생생물 기수갈고둥, 붉은발말똥게 등은 2024년 조사에서 추가로 확인되었다. 이는 조사 강도 증가의 영향일 수 있으나, 그럼에도 주교천하구습지가 기수성 생물의 주요 서식처로 기능하고 있음을 시사한다.
4. 고 찰
4.1 식생 분포에 따른 습지 생태계 특성
주교천하구습지의 대부분 구간에서 석축으로 보강된 인공제방이 축조되는 등 직강화가 진행되었음에도 하도의 곡류 형태가 비교적 잘 보전되어 사력퇴, 하중도 등 다양한 퇴적 지형 발달이 습지식생 정착에 긍정적으로 작용하고 있었다. 그러나 완경사 홍수터의 부재, 콘크리트 제방 및 논둑 조성 등 물리적 제약으로 인해 전체 18개 식물군락 중 대상식생, 경작지식생 등 육상식물군락이 12개(66%)를 차지하였다. 수생 및 습생식물군락은 5개로, 거제시부터 남해군까지 남해안 일대 하구습지에서 보고된 6 - 12개 군락 구성과 비교하면 식생 다양성은 다소 낮은 수준으로 판단된다(NIE 2022). 그럼에도 불구하고 수변식생평가지수(KERVI)는 B등급(70점, ‘좋음’)으로 평가되었으며, 세부 분석 결과 외래식물군락과 육상식물군락의 분포 면적 비율이 상대적으로 낮아 습지생태계의 건전성이 양호한 것으로 나타났다(NIE 2024). 일정한 수위 및 유속이 유지됨에 따라 발달한 퇴적 지형을 중심으로 갈대군락의 고밀도 단일 우점과 제방에서 하도로 곧바로 이어지는 지형 구조로 인한 홍수터 발달의 제한이 복합적으로 작용하여 외래식물 및 육상식물군락의 침입을 억제한 것으로 판단된다(van der Putten 1997, Richardson et al. 2007).
본 연구 지역을 대표하는 단위 식생은 갈대군락으로, 하구습지 전역에 걸쳐 퇴적지형이 발달한 지역에서는 예외 없이 밀도 높게 분포하며 전체 식생 면적의 약 75%를 차지하였다. 이러한 대규모 군락 형성은 평균 수심 10 cm, 경사 1° 미만의 완경사 등 주교천의 지형적 특성과 수리·수문 조건이 갈대 생육에 적합한 환경을 제공하기 때문으로 판단되다(NIE 2024, Lee et al. 2025). 또한 갈대는 과습한 환경과 혐기성 토양에서의 생존 능력, 0-22.5% 범위의 염분에 대한 높은 내성 등 넓은 생태적 지위와 경쟁력 있는 생리적 특성을 보이기에 해당 환경에서 우점종으로 정착하고 군락을 안정적으로 유지할 수 있는 것으로 보인다(Lee 2020).
이러한 갈대군락의 단일 우점 특성으로 인해 일반적으로 하구습지에서 출현하는 갈풀군락(Phalaris arundinacea)과 줄군락(Zizania latifolia)은 본 조사에서 확인되지 않았다. 해당 군락은 유수역 또는 정수역 주변의 잦은 범람을 경험하는 홍수터를 선호하는 것으로 알려져 있으나, 주교천하구습지는 수변부와 제방이 곧바로 접하는 구조적 특성으로 인해 홍수터가 충분히 형성되지 못하는 지형적 제약과 더불어, 해수의 유입에 따른 기수 환경의 염분 조건 역시 이들 식생에게 불리하게 작용한 것으로 판단된다(Tang et al. 2022, Delaney et al. 2025). 한편, 1979년과 1996년에 촬영된 항공사진 분석 결과, 직강화가 완료된 이후 시점임에도 현재와 유사한 곡류하도 및 퇴적 지형이 당시에도 형성되어 있었던 것으로 확인되며(NGII 2025), 이는 하천 정비 이후에도 일부 구간에서 자연적인 하도 발달이 지속되어 왔음을 시사한다(Fig. 4). 이러한 지형적 안정성은 저지대 습지 형성과 더불어 갈대군락의 장기적인 유지에 물리적 기반을 제공해 온 것으로 추정된다. 특히 한 번 형성된 갈대군락은 지하경 확산, 고밀도 피복 등에 의해 타 식물의 유입을 제한하는 생태적 특성을 보이므로, 이러한 요소들도 현 군락 구조의 고착화에 기여했을 것으로 판단된다(Amsberry et al. 2000, Weis and Weis 2003). 다만, 항공사진 분석은 토지피복 및 하도 형태 변화를 정성적으로 파악하는 수준에 그쳐, 식생의 분포 범위나 종조성 변화를 정량적으로 해석하는 데에는 한계가 있다. 이에 따라 향후에는 과거 식생조사, 수문자료, 지형고도자료 등을 통합한 다원적 분석을 통해 보다 정량적인 접근이 필요할 것이다.

4.2 생물종 분포 특성
주교천하구습지 내 곡류부와 합류부를 중심으로 사력퇴, 포인트바, 간석지 등 다양한 퇴적지형이 밀집되어 분포하고 있으며, 점토에서 거력에 이르는 다양한 입경의 하상구조가 관찰되었다(NIE 2024). 이러한 복잡한 지형 구조와 완만한 하천 구배는 해수의 유입을 용이하게 하여 기수역 특성을 형성하고, 하천성과 해양성 퇴적물이 함께 집적될 수 있는 물리적 기반을 제공한다. 결과적으로 이들은 다양한 생물종의 생태적 특성에 부합하는 미소서식처가 형성됨에 따라 생물다양성 유지 및 증진에 기여하고 있는 것으로 판단된다. 따라서 주교천하구습지에는 총 1,074종의 생물이 확인되었으며, 담수, 기수, 해양 환경을 모두 아우르는 다양한 기능군의 생물군이 포함되었다. 습지 내 구간별로 염분과 토지이용 조건에 따라 분류군별 출현 양상에는 다소 차이가 있었다. Ⅲ구간의 염도는 0.1 – 0.7%로 해수의 영향을 거의 받지 않는 담수역에 해당하여 담수무척추동물 및 1차 담수어류가 주로 확인되었다. Ⅱ구간은 염도가 0.5 - 10.9%로 급격한 염분 변화가 관찰되는 과도 구간으로 담수 및 기수 무척추동물이 공존할 수 있었으며, 주변 농경지는 조류의 풍부도에 긍정적으로 작용하고 있었다. Ⅰ구간은 섬진강 본류로 해수가 직접 유입되어, 기수무척추동물, 해양성 어류 및 철새 도래지 역할로 조류의 풍부도가 상대적으로 높게 나타났다.
하구습지의 특성을 가장 잘 반영하는 분류군은 기수역의 염분 환경에 직접적인 영향을 받는 어류와 저서성대형무척추동물이다(Basset et al. 2013). 어류의 경우, 경제적 가치는 낮지만 먹이망 구조 상 중요한 위치를 차지하고 있는 망둑어과 어종(11종, 885개체)의 우점, 열린 하구 특성으로 인해 상류역(Ⅲ구간)까지 회유성 어종(은어, 숭어 등)이 도달하는 점, 2021년도 미기록종으로 보고된 흰점실고기의 출현, 그리고 생태계교란 생물인 블루길과 배스의 미출현 등의 요소가 복합적으로 작용하였다. 이러한 특성에 따라 하구어류평가지수(KEFAI)는 ‘매우 좋음’에 해당하는 A등급(82.7점)으로 평가되었으며, 본 지역의 어류 군집은 높은 생태적 건강성을 유지하고 있는 것으로 판단된다. 또한 다양한 입경의 하상기질로 구성된 염습지와 대규모 갈대군락의 발달이 서로 맞물려 말똥게, 방게, 기수우렁이 등 기수성 무척추동물이 대규모로 서식할 수 있는 풍부한 먹이원과 은신처를 제공하고 있었다. 이로 인해 생산성이 높은 하구 생태계가 유지되고 있으며, 하구저서성 대형무척추동물평가지수(KEBMI)는 ‘좋음’ 수준인 B등급(71.4점)으로 평가되었다. 특히 멸종위기 야생생물인 붉은발말똥게, 흰발농게의 개체군이 일정 규모 이상 유지되고 있는 점은 본 하구습지의 보전 가치를 높이는 생태적 지표로 판단된다.
주교천하구습지의 특징 중 하나인 갈대군락의 발달은 설치류와 같은 소형 포유류에게도 서식처 및 은신처로 기능하고 있으며, 이는 다시 삵, 너구리 등 중대형 포식성 포유류의 출현으로 이어져 상위 영양 단계가 유지되는 안정적인 생태계 구조 형성에 기여하고 있는 것으로 판단된다(Estes et al. 2011, Poulin et al. 2002). 특히 수환경에서 최상위 포식자로 알려진 수달은 가족 단위의 개체군이 확인되어 본 지역에서의 안정적인 서식이 이루어지고 있음을 시사하며, 핵심종으로 기능할 가능성이 높은 종으로 평가된다. 포유류 뿐만 아니라 육상곤충에게도 갈대, 억새, 대나무 등의 식물군락은 먹이원과 은신처로서 기능하고 있어, 700여종에 달하는 높은 종다양성이 관찰된 것으로 판단된다(Landis et al. 2000). 특히, 하구습지는 비행성을 지진 곤충에게는 확산 및 소멸 동태에 영향을 미치는 중요한 매개지점이 되며, 곤충상 중에 기후변화 지표종, 생태 정보가 부족한 미기록종 등이 포함되어 있어 곤충 생태 및 생물지리학적 연구 측면에도 학술적 가치가 높을 것이다(Bilton et al. 2001).
하천 주변 논습지 및 섬진강 합수부에 발달한 염습지 등 다양한 서식 공간은 계절적 서식처 및 먹이터로 기능하여, 노랑부리백로, 큰고니, 큰기러기 등 멸종위기 야생생물 포함 다양한 수조류가 관찰되었다. 특히 주교천하구습지를 중심으로 광양만, 갈사만, 섬진강 하구 등 인근 주요 철새도래지와의 생태적 연결성이 확보되어 있어, 이 일대를 중심으로 연간 30,000개체 이상, 멸종위기 야생생물 15종 포함 총 87종의 조류가 확인된 것으로 보고되었다(GREF 2022, NIER 2024). 이러한 결과는 주교천하구습지가 섬진강 하구, 광양만과 연계된 배후 서식지로 기능하고 있으며, 큰기러기, 노랑부리백로 등 국제적으로 중요한 희귀조류의 중간 기착지 또는 서식처로서 높은 보전 가치를 지닌다는 점을 시사한다. 한편 본류구간에서는 기수역이 형성되어 대부분의 양서·파충류에게는 서식 스트레스 요인으로 작용함에 따라 고유종인 한국산개구리 포함 큰산개구리, 남방도롱뇽 등의 양서류는 제내지에 위치한 농경지, 둠벙, 농수로 등 수환경을 중심으로 번식이 확인되었다(Hopkins and Brodie Jr 2015). 생태계교란 생물로 지정된 황소개구리와 붉은귀거북은 조사구역 전반에 걸쳐 광범위하게 분포하고 있으며, 제방도로를 따라 파충류의 로드킬도 빈번하게 발생하고 있어 지속적인 관리 및 서식지 연결성 확보가 필요할 것으로 판단된다.
최근 5년간 수행된 동천하구, 한강하구 등 국내 주요 6개 하구습지의 정밀조사 결과, 평균 종 수는 멸종위기 야생생물 13종 포함 912종으로, 이와 비교하면 주교천하구습지의 생물다양성은 이를 상회하는 수준으로 확인되었다(NIE 2024). 갈대군락, 염습지, 논습지 등 다양한 서식처의 공존과 개방된 하구 특성은 생물종의 유입과 확산을 용이하게 하여 겨울철새, 기수성 생물에게 적합한 서식 환경을 제공하는 것으로 판단되었다. 또한 주교천하구습지는 섬진강 하구 및 광양만과 직접적으로 연계되어 있어, 향후 체계적인 생물다양성 보전이 이루어진다면 하천-하구-연안까지 이어지는 습지생태축의 연속성 확보 및 연결성 강화를 통해 국가 차원의 생물다양성 관리에도 긍정적인 영향을 미칠 수 있을 것으로 기대된다. 따라서, 본 연구 결과는 주교천하구습지의 보호지역 지정 가능성을 과학적으로 뒷받침할 수 있는 실질적 근거자료로 활용될 수 있을 것이다.
5. 결 론
주교천하구습지는 섬진강과 광양만 사이에 위치한 하구습지로서, 하천–하구–연안으로 이어지는 남해안 생태축 상의 전략적 연결지점에 해당한다. 조사 결과, 멸종위기 야생생물을 포함한 핵심종의 안정적인 서식이 확인되었으며 기수역에 특화된 다양한 생물군집의 출현 또한 뚜렷하게 나타났다. 특히 하구습지 전반에 걸쳐 광범위하게 분포하는 갈대군락은 수질 정화 뿐만 아니라 조류와 기수성 무척추동물의 산란·은신처로 기능하여, 외부 교란을 저감하는 생태적 완충지대로서의 역할을 수행하고 있었다. 이러한 결과는 주교천하구습지가 국내는 물론 국제적인 관점에서도 생물다양성 보전 가치가 높은 습지임을 시사한다.
조사지역의 주요 구간은 과거 하천정비로 인해 직강화된 인공적 구조를 지니고 있음에도, 곡류하도, 사력퇴, 하중도 등 다양한 퇴적지형이 장기적으로 유지되고 있어 미소서식처의 발달 및 습지식생 정착에 기여하고 있었다. 식물군락의 구조적·기능적 다양성은 다소 낮게 나타났으나, 하천 단면 구조의 자연화, 경사 완화, 자연형 홍수터의 재조성 등 물리적 조건의 개선이 이루어진다면 습지가 지닌 고유의 회복 탄력성을 바탕으로 식물군락의 다양성은 점진적으로 향상될 것으로 판단된다. 이외에도 중상류부 농경지 및 유수지로부터의 비점오염원 유입, 중금속(납, 카드뮴, 수은) 검출, 하류부 골프장 운영 등은 생태계 안정성을 저해하는 요인으로 작용하고 있어, 중금속 검출 구간에 대한 정밀 오염도 평가와 복원 우선구간 설정, 농경지 및 골프장 주변의 완충지대 조성, 주기적인 수질 모니터링 체계 도입 등 통합감시 기반의 생태관리체계 구축이 요구된다.
한편, 조사구간에서는 총 10종(식물 5종, 양서·파충류 2종, 육상곤충 3종)의 생태계교란 생물이 확인되었으며, 이 중 환삼덩굴, 단풍잎돼지풀은 하천 제방부를 중심으로 확산되고 있었다. 특히 하천정비 이후 장기간 방치된 제방과 둔치부는 외래식물의 확산 거점이자 주요 경로로 기능하고 있어, 자연 식생의 회복을 유도하기 위해서는 외래식물 우선관리 구역 설정 및 계절별 제거계획 수립, 토착종 중심의 식생 복원과 모니터링 기반의 사후관리 체계를 병행해야 할 것이다. 더불어 본 지역은 고포수변공원, 갈사만갯벌, 남파랑길 등과 연계된 광역 생태·문화 네트워크의 거점으로서의 잠재력이 높으며, 향후 생태교육, 탐방 프로그램, 지역주민 참여형 보전사업을 통해 지속가능한 생태관리 모델로 발전할 수 있을 것이다. 이는 단지 특정 습지의 보전을 넘어 하구습지 생태계의 회복과 연안 통합관리라는 정책적·사회적 차원에서도 중요한 함의를 지닌다.
다만 본 연구는 단년도 조사에 기반하고 있어, 계절 변화나 연차 간 생물상 변동, 장기적인 서식처의 구조적 변화 등을 충분히 반영하지 못한 한계가 있었다. 따라서 향후에는 주요 생물군집의 생태적 반응을 시간 축에서 추적할 수 있는 장기 생태 모니터링 체계를 구축하고, 누적 자료를 활용한 정량적 평가와 우선관리 구간의 재설정을 통해 대상지의 보전 및 관리 전략을 지속적으로 보완해 나갈 필요가 있을 것이다.
