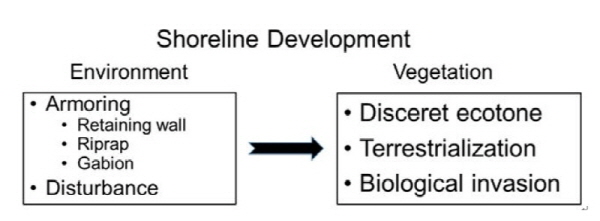
1. 서 론
2. 연구 방법
2.1 조사지 개황
2.2 현장 조사
2.2.1 호안 구조
2.2.2 호안 식생
2.3 자료처리
3. 결과
3.1 호안의 물리적 구조
3.2 호안 식생
4. 고찰
1. 서 론
하천을 댐으로 막아서 조성한 저수지는 인위적으로 조절되는 수위 변동의 범위에서 새로운 호안 (湖岸, shoreline; 본 본문에서는 혼돈을 피하기 위하여 護岸은 보강 호안으로 표현함)이 발달한다 (Nilsson and Berggren 2000). 호안의 어원적 정의는 육상과 물의 물리적 경계인 1차원의 선이다. 그러나 호수, 저수지 및 하천에서 수위가 역동적으로 변동하므로 호안의 범위는 시간에 따라서 변화하고 (Boak and Turner 2005), 물이나 육상의 상호작용이 영향을 미치는 공간이므로 1차원이 아닌 2차원 면으로 파악될 수 있다. 더욱이 호안에는 식생이 발달하므로 물, 토양, 대기 및 생물을 포함하는 3차원 공간으로서 호안을 규정할 수 있다. 결국 호안은 육상 생태계와 수생 생태계가 연결된 지역으로서 직접적이고 강력한 상호작용에 의하여 이 두 생태계가 밀접하게 연결되어 있다. 이러한 호안은 다양한 생물군집이 발달한 생물다양성의 보고일 뿐만 아니라 생지화학 기능의 핵심공간이다 (Strayer and Finlay 2010).
북한강 수계에 조성된 춘천호, 의암호, 청평호, 팔당호 등과 같이 발전용 댐호의 호안에서는 댐 조성 전에 존재하던 육상 식물이 파괴되고 새로이 식생이 정착하여 있다. 이들 저수지는 연중 수위가 안정되어 있어서 수생식물이 생육하는 연안대 (littoral zone)가 발달할 수 있다 (Cho and Cho 2013). 호안 식생은 침식을 방지하고 제방을 안정화시키고, 조류 성장을 유발하는 영양소를 제거하고, 쇄설물과 오염물질을 여과하고, 야생생물의 먹이와 은신처를 제공하고, 수체에 그늘을 드리우고, 풍력과 파력을 감세하고, 심미적 가치를 제공하는 기능을 한다 (Elias and Meyer 2003). 그러나 호안 생태계는 교란에 민감하고 인간의 정주와 휴양 등의 여러 활동에 적합한 장소로서 (Liddle and Scorgie 1980, Ostendorp et al. 2004, Schmieder 2004), 호수와 저수지의 호안은 인간의 간섭에 의하여 심하게 변형되어 왔다 (Brauns et al. 2011). 호안과 이와 가까운 지역에서는 토지 이용이 호안 식물의 풍부도에 영향을 주는 주요한 요인으로서 (Dodson 2008, Alahunta et al. 2012), 특히 물의 사용, 호안 구조의 변화, 도시 개발 등은 수변 및 연안대의 수생식물 군집에 심각한 손상을 입힌다 (Schmieder 2004, Jeppesen et al. 2011).
산업화와 도시화가 진행됨에 따라서 북한강 수계의 호수에서 호안 정비, 수변부 개발 등의 지속적인 교란으로 인해 수변식물의 생육지가 제한되고 식물상과 식생구조의 변화가 초래되고 있다. 본 연구의 조사지인 의암호는 강원도 춘천시의 북한강 수계에 위치한 인공호수로서 수력발전을 목적으로 북한강과 소양강을 막는 의암댐의 축조에 의하여 생겨났다. 의암호는 연간 수위변동 폭이 1 m 내외로서 좁고 수변부의 경사가 완만하여 호안식생이 생육하기에 적합한 환경이다 (Cho and Cho 2013). 그러나 특히 최근 보강 호안의 축조, 자전거 도로 건설, 수변부 공원 조성 등으로 호안 환경에 교란이 심하게 일어나고 있다. 따라서 본 연구에서는 북한강 수계에 위치한 인공호인 의암호에서 수변 환경의 변화에 따른 호안 식생의 변화를 파악하기 위하여, 1) 수변의 호안 유형을 구분하고 분포 특성을 조사하고, 2) 2006년, 2010년 및 2013년에 조사된 호안 식생의 구조와 분포의 변화 특성을 분석하여, 3) 수변부 교란에 따른 호안 식생구조의 변화를 파악하는데 연구의 목적이 있다.
2. 연구 방법
2.1 조사지 개황
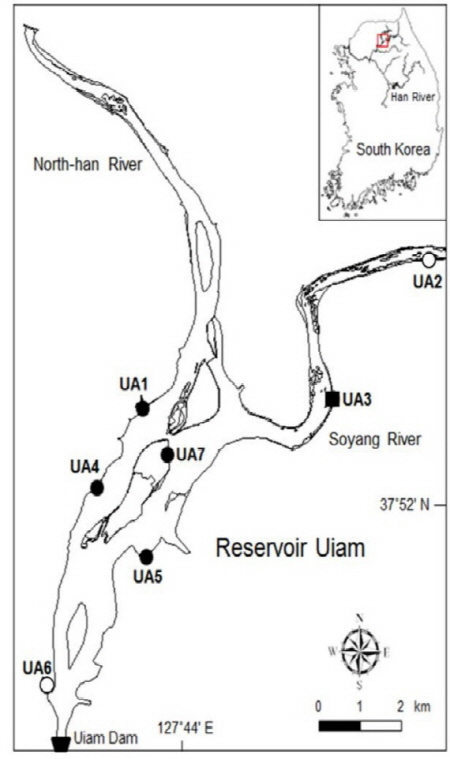
본 연구의 조사지인 의암호는 강원도 춘천시에 위치한 인공호로서 북한강에 수력발전소를 만들기 위한 의암댐을 1967년에 축조하면서 형성되었다. 의암호는 북한강의 춘천댐과 소양강의 소양강댐으로부터 하천수가 유입되고 있다. 이 저수지의 형태는 하천 형상이었고 수면적은 15.0 km2, 유역 면적은 7,709 km2이다 (WAMIS 2015). 또한 총저수용량은 8,000x107 m3, 평균 수심 5.3 m, 평균 호안 경사 2.6˚, 연수위변동폭 1.6 m yr-1으로서 수위가 안정적으로 유지되고 호안이 완만하여 호안식생이 발달할 수 있는 환경이었다 (Cho and Cho 2013). 한편 소양강댐 하류의 소양강 유입부와 춘천댐 하류의 북한강 유입부는 거의 하천의 하안 환경을 나타내고 있었다. 의암호 호내에는 위도, 상중도, 하중도, 붕어섬 등의 섬이 다수 위치하였다. 본 저수지는 춘천 도심에 인접하여 휴양 목적으로서 이용되고 있어서 인간 간섭에 의한 교란에 매우 취약한 상황이다.
의암호에서 호안 식생의 조사지점은 수생식물이 생육하는 연안대가 발달된 곳, 다양한 호안 식물이 분포하는 곳, 전체 호소의 호안 식생을 대표할 수 있는 곳을 중심으로 선정하였다 (Fig. 1). 선정된 조사지점으로서 강원도 춘천시 서면에 위치한 의암호 서쪽 수변의 3개 조사지점 (UA-1, UA-4 및 UA-6), 동면에 위치한 소양강 유입부의 2개 조사지점 (UA-2와 UA-3) 및 삼천동의 공지천 유입부 인근 조사지점 (UA-5)은 2006년, 2010년 및 2013년에 조사하였다. 또한 강원도 춘천시 중도동에 위치한 호안 섬의 1개 조사지점은 2013년에 새로이 조사지로 선정하였다. 각 조사지점에서 호안 식생 조사는 홍수 전 하계인 6월말과 7월초에 실시하였고, 식생도 작도는 2010년과 2013에 홍수 후 추계에 실시하였다. 2006년과 2013년 현장 조사는 본 연구자에 의하여 수행되었고 (HERC 2007, 2014), 2010년 조사 결과는 HERC (2011)를 이용하였다. 한편 호안의 물리적 구조와 개수 유형은 2015년에 조사하였다.
|
Fig. 1. Map showing the study area in the Reservoir Uiam (closed circle, sampling site of the riprap shoreline; closed square, the gabion shoreline; open circle, the soil shoreline). |
2.2 현장 조사
2.2.1 호안 구조
의암호의 전체 수변에서 호안의 물리적 구조 및 개수 현황을 파악하였다. 2015년 5월에 차량과 선박을 이용하여 평수위에서 호안 유형을 조사, 분류하였다. 호안 유형은 크게 자연 호안과 개수 호안으로 분류하였고, 자연 호안은 다시 흙 사면 (soil slope)과 암벽 (rock face)으로 개수 호안은 돌망태 (gabion), 사석 (riprap), 옹벽 (retaining wall)으로 다시 세분하였다 (Fig. 2). 돌망태는 철망 안에 돌을 채운 것이고, 사석은 채석장에서 크게 쪼개진 돌을 쌓았고, 옹벽은 콘크리트 블록을 급경사로 쌓아 올린 구조이다.
|
Fig. 2. Photographs showing the shoreline types in the Reservoir Uiam. (a) soil slope, (b) rock face, (c) riprap, (d) gabion and (e) retaining wall. |

2.2.2 호안 식생
선정된 의암호 조사지점의 호안에서 만수위 높이부터 저수지의 유심선 (thalweg)을 향하여 트랜섹트를 설치하였다. 트랜섹트 상에서 이동하면서 식물 군집을 구분하고 줄자 또는 레이저 거리측정기를 이용하여 각 식물 군집의 거리를 측정하였다. 트랜섹트에서 나타나는 식물 군집에서 5 m x 5 m 넓이의 방형구를 설치하고 출현 식물종과 종별 피도를 기록하였다. 출현 식물의 동정과 명명은 Lee (1993), Park (1995) 및 Choi (2000)를 따랐다. 출현종의 습생도 (수생식물, 습생식물 및 중건생식물) 및 수생식물 생장형 (침수식물, 부유식물, 부엽식물 및 정수식물)의 구분은 Choung et al. (2012)를 따랐다.
식생도는 다음 지도 (http://map.daum.net/)의 항공사진을 바탕지도로 하여 수생식물이 나타나는 지역을 현장을 답사하면서 식생 분포 확인하고 분포하는 식물 군집명을 기록하였다. 필요에 따라서 레이저 거리 측정기로 거리를 측정하였다. 식물 군집명은 우점종에 의하여 이름을 붙였다. 현장에서 작도한 식생도를 QGIS (QGIS Development Team 2015)를 이용하여 현존 식생도를 작도한 후 식물군집 별 분포면적을 산출하였다.
2.3 자료처리
각 조사지점별 식생 자료를 구축하기 위하여 트랜섹트에서 조사한 식물 군집의 종별 피도 자료를 각 군집이 차지하는 거리를 가중치로 하여 조사지점에서 식물종별 평균 피도를 산출하였다. 이 식생 자료를 이용하여 습지지수 및 다변량 분석을 실시하였다.
조사지점에서 습지 구조를 어느 정도 반영하는지를 평가하기 위하여 다음 식에 따라서 조사지 습지지수 (site wetland index, SWI)를 구하였다 (Coles-Ritchie et al. 2007, Son et al. 2015).
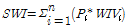 (Eq. 1)
(Eq. 1)
여기에서 Pi는 전체 식물종의 피도 총계에 대한 식물종 i의 평균 피도 비율이다. 또한 WIVi는 식물종 i의 습지지표지수 (wetland indicator index)이다. 습지지표지수는 절대육상 지표식물이 1, 임의육상 지표식물이 25, 양생 지표식물이 50, 임의습지 지표식물이 75, 절대습지 지표식물이 100을 나타낸다. 조사지 습지지수 (SWI)가 100에 가까운 값일수록 습지로 유지되고 있으며, 1에 가까울수록 육상화가 이루어졌음을 의미한다.
의암호의 조사지점에서 식생구조를 파악하기 위하여 다변량 분석을 실시하였다. 3개 조사연도 (2006년, 2010년 및 2013년)의 식생 자료에서 피도 합이 3% 미만인 종과 출현 빈도가 3회 미만인 종은 제외하였다. 준비된 식생 자료를 이용하여 R 환경 (R Development Core Team 2013)에서 Vegan package (Oksanen et al. 2015)의 ‘decorana’ 함수를 이용하여 탈경향대응 분석 (detrended correspondence analysis, DCA)을 실시하였다.
3. 결과
3.1 호안의 물리적 구조
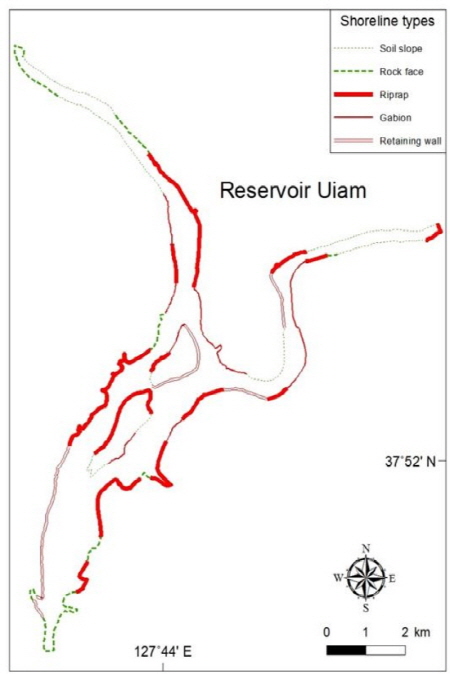
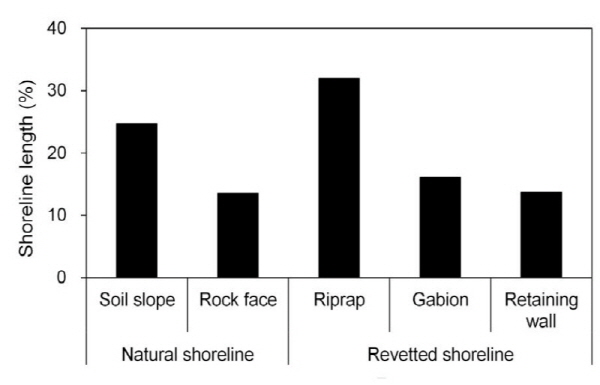
의암호는 산지와 평지를 지나는 북한강과 소양강에 형성되어 자연적인 호안은 경사가 완만한 곳에서 흙 사면과 경사가 급한 곳에서 암벽이다. 그러나 호안에 다양한 인위적인 개수가 이루어지면서 이러한 자연 호안이 인위적인 형태로 변화하였다 (Fig. 3). 자연 호안인 흙 사면과 암벽은 주로 춘천댐과 소양강댐 하류 및 의암댐 직상류에 잔존하고 있었다. 이를 제외한 의암호의 중심부는 대부분 개수 호안이었는데, 콘크리트 블록의 옹벽은 의암댐 인근 서안, 상중도 동안 및 소양강 유입부에 위치하였고, 나머지 호안이 사석과 돌망태로 축조되어 있었다 (Fig. 3). 의암호 전체 호안에서 자연 호안이 약 40%, 개수 호안이 약 60% 이었고, 개수 호안 중에서 사석 > 돌망태 > 옹벽의 순서로 많았다 (Fig. 4).
3.2 호안 식생
의암호 호안의 7개 조사지점에서 2006, 2010 및 2013에 조사한 식생 자료를 이용한 탈경향대응 분석 (DCA)에서 제1축과 제2축이 총 변이의 각각 42.5%와 27.9%를 설명하였다 (Fig. 5). 먼저 출현 식물종의 배열에서 하천 홍수터에 주로 분포하는 달뿌리풀 (Phra.jap), 갯버들 (Sali.gra), 갈풀 (Phal.aru) 등은 배열에서 왼쪽에 위치하였다 (Fig. 5a). 나도겨풀 (Leer.jap), 송이고랭이 (Scir.tri), 세모고랭이 (Scir.trq), 물꼬챙이골 (Eleo.mam) 등은 주로 수심이 얕은 습지에 출현하는 종으로서 그림의 오른쪽에 배열하였다. 나머지 줄 (Ziza.lat), 큰고랭이 (Scir.lac), 갈대 (Phra.com), 애기부들 (Typh.ang), 개발나물 (Sium.sua) 등의 수생식물과 쑥 (Arte.pri), 가시박 (Sicy.ang), 단풍잎돼지풀 (Ambr.tri), 족제비싸리 (Amor.fru) 등의 외래종을 포함한 중건생식물이 중앙에 섞여 있었다.
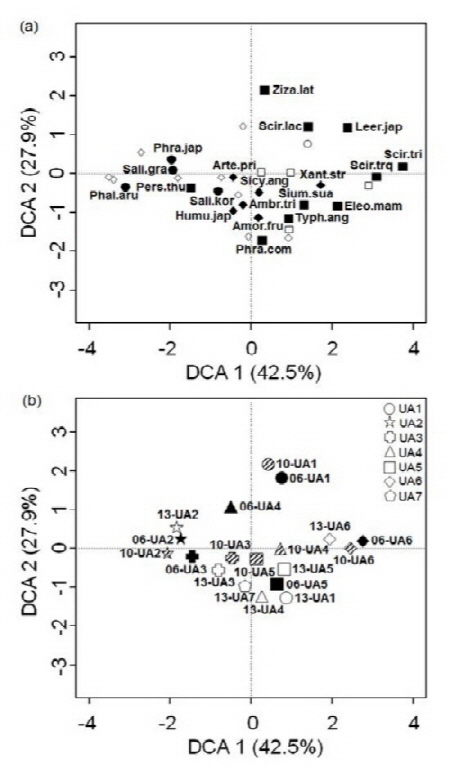
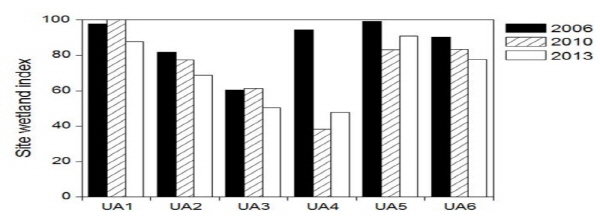
DCA 조사지점의 배열에서 소양강 유입부인 흙 사면 호안의 조사지점 UA2와 돌망태 호안인 UA3는 그래프의 왼쪽에 위치하였고 의암호 중앙에 위치한 사석 호안의 조사지점 UA4, UA5 및 UA7은 그래프의 중앙에 위치하였으며 소하천 유입부에 위치한 흙 사면의 조사지점 UA6는 그래프의 오른쪽에 위치하였다 (Fig. 5b). 이상의 결과를 종합하면 소양강 유입부의 조사지점 UA2와 UA3은 하천 하안식생 유형으로 구분되었으며 특히 흙 사면인 UA2보다 돌망태 호안인 UA3가 더 중앙에 위치하여 UA3가 중건생식물과 수생식물의 중요도가 높아지는 것으로 나타났다. 또한 의암호의 중앙부에 위치한 조사지점 중에서 UA1, UA4, UA5, UA6 및 UA7 중에서 흙 사면인 UA6이 얕은 수심에서 생육하는 수생식물이 발달하였고 사석 호안인 다른 조사지점에서는 수생식물과 중건생식물의 혼재된 식생구조를 보이고 있었다. 따라서 돌망태 및 사석 호안에서 호안의 환경 경사가 완만하지 않고 수생식물과 중건생식물의 생육지가 중간 전이지역이 발달하지 않고 단절되어 나타났다. 실제로 사석 호안에서는 물과 육상의 접경지역에 거석을 쌓아서 물과 육상역이 부드럽게 연결되지 못하고 단절되는 지형적 특성이 나타났다. 각 조사지점에서 시간에 따른 식생구조의 변화를 살펴보면, DCA 결과에서 2006년부터 2013년까지 시간이 경과함에 따라서 바깥쪽에서 중앙부로 이동하는 경향이었다. 앞에서 지적했듯이 중앙으로 이동한다는 것은 시간이 경과함에 따라서 수생역에서 육상역으로의 연속성에 단절이 심화되고 있다는 것을 나타낸다. 이러한 결과로서 모든 조사지점에서 조사지 습지지수 (SWI)는 2006년부터 시간이 경과함에 따라서 대체로 감소하는 경향이었다 (Fig. 6).
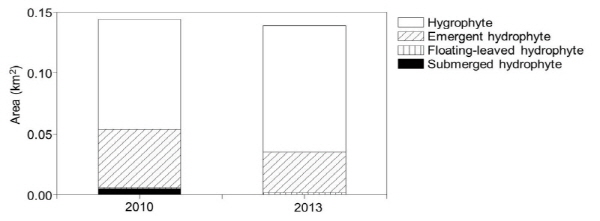
의암호의 각 조사지점 주변에서 조사한 식생 분포면적은 2010년의 0.145 km2에서 2013년에 0.139 km2로서 다소 감소하였다 (Fig. 7). 이중에서 2013년에 수생식물 분포면적은 감소한 반면에 습생식물의 분포면적은 다소 증가하였다. 특히 수생식물 분포면적 중에서 침수식물의 분포면적이 크게 감소하였다.
4. 고찰
북한강 수계에 위치한 발전용 저수지인 의암호는 수변부의 경사가 완만하고 수위가 안정적으로 유지되며 연안대가 발달할 수 있는 지형이 많이 존재하기 때문에 수변식생이 생육하기에 적합한 환경이다. 최근 수변을 따라서 사석쌓기 자전거 도로 건설, 공원 조성 등이 시행되어 수변 환경이 변화하였다. 이러한 인간 요인에 의한 수변부 경관의 변화가 자연적인 일차천이보다 영향력이 더 크다고 알려져 있다 (Décamps et al. 1988).
의암호에서 댐으로부터 상중도까지의 호안은 수변부의 폭이 좁고 경사가 급하며 인공제방으로 되어있다 (Fig. 3). 반면에 춘천댐과 소양댐 하류에서는 대부분의 호안이 경사가 완만하고 폭이 넓게 발달한 자연적인 지형으로서 갈대, 달뿌리풀, 버드나무 군락이 넓게 우점하고 있다. 인공제방 중 거석 사석이 존재하는 부분에는 식생이 거의 존재하지 않았다. 특히 2010년부터 시행된 자전거 도로 건설로 인하여 서쪽 호안의 급경사 호안에 옹벽이 설치되어 식물의 생육 가능지가 크게 훼손되었다. 따라서 최근 의암호에서는 호안 정비에 의하여 호안식물이 정착이 가능한 생육지가 크게 감소되어 호안 생태계의 자연성이 훼손된 것으로 판단되었다.
의암호의 수변부에서 중건생식물의 출현이 지속적으로 증가하였는데 (Fig. 5), 이는 의암호의 수변부가 중건생식물이 생육하기 적합한 보다 건조한 환경으로 변하고 있다는 것을 반영한다. 이러한 육역화 현상은 의암호의 조사지점에서 습지지수와 분포면적 조사 결과에서도 확인할 수 있었다. 조사지 습지지수는 습지화 정도를 비교할 수 있는 유용한 지표인데 (Cole-Ritchie et al. 2007), 의암호에서 최근 습지지수가 점차 감소하여 양생지역에 가까워졌으며 특히 서쪽 호안에서 급격하게 감소하였다 (Fig. 6). 이러한 결과는 서쪽 호안이 더 잦은 수변부 개발에 의하여 수변부의 연속적인 환경 변화가 파괴되었기 때문이라고 생각된다. 또한 조사지점에서 최근 수변 식생의 총분포면적이 감소하고 있으며 특히 수생식물의 분포면적은 감소한 반면 습생식물의 분포면적은 다소 증가하였다 (Fig. 7). 이러한 결과는 수변부가 인위적인 교란으로 인해 생육지가 점점 감소하고 있으며, 육상화가 진행되어 수생식물의 분포지가 습생식물의 분포지로 대체되고 있기 때문으로 판단된다.
의암호에서 생태계교란식물에 속하는 외래종인 가시박, 단풍잎돼지풀, 돼지풀 등이 대부분의 조사지점에서 출현하였다. 수변지역에서 외래종 침입은 일반적으로 인간 활동에 의한 서식처 환경 조건의 악화와 밀접히 관련되어 있으며 이로 인하여 고유종의 경쟁력을 감소시킨다 (Thiebaut et al. 2007). 한편 의암호 서쪽 호안에 위치한 조사지점에서는 갈대와 족제비싸리 우점 군집으로 지속적으로 변화하는 경향을 보였다 (Fig. 5). 특히 이곳에서는 자전거 도로 건설, 호안 정비 등의 수변 공사가 빈번히 진행되어 호안에 교란이 심한 곳이었다. 일반적으로 호안에서 교란은 수변부에서 인간의 토지이용과 깊은 관계를 가진다 (Corbacho et al. 2003, Jennings et al. 2003).
이상의 결과를 종합하면, 춘천시에 위치한 의암호에서는 최근 옹벽, 사석쌓기, 돌망태에 의한 호안 보강과 낚시, 수상 레져, 관광 등의 각종 교란이 호안에 가해지고 있다. 이에 따라서, 호안 식생은 수생식물과 육상식물이 연결되는 점이대 (ecotone)가 불연속적으로 단절되고 있으며, 연안대 식생이 점차 육역화되어 습생 및 육상 식생으로 변하고 있으며, 교란된 식생에는 외래식물이 빈번히 침입하고 있다고 판단된다 (Fig. 8). 일반적으로 저수지 호안에서 식물 군집의 성립을 결정하는 중요한 요인은 경사와 노출 (exposure)이다 (Luken and Bezold 2000). 토양이 잔존하고 경사 완만한 호안에서는 습지 식물종이 풍부하고 보전 가치가 높은 종이 많다 (Luken and Bezold 2000). 그런데 자연적인 완만한 경사의 호안에 각종 보강 호안이 조성되면 이로 인하여 수변부가 급격히 단절되는 결과를 가져오고 이에 따라서 생물다양성이 높은 호안식생이 발달이 어렵게 된다. 미국의 Chesapeake Bay에서 실제로 침수식물의 분포가 사석 호안에 의하여 심하게 제한되고 있었다 (Patrick et al. 2014). 또한 보강 호안과 교란에 의하여 개발된 호안이 미개발된 호안보다 총종수와 자생종수가 적었고 수생식물 군집에서 급하게 육상식물 군집으로 변화하는 특성을 보였다 (Elias and Meyer 2003). 결론적으로 의암호에서 호안 정비와 수변부 개발에 의하여 수변 식생의 생육지가 제한될 뿐만 아니라, 육역화에 의하여 수역부터 육역까지 횡적 연속성이 손실되어 육역화된 식생의 출현이 증가하고 있었다. 따라서, 의암호에서는 호안 생태계의 보전을 위하여 불필요한 보강 호안의 조성을 억제하고, 토양 침식의 위험성이 낮은 곳에서는 조성된 보강 호안을 해체하거나 재조성하여 호안의 복원을 추진하여야 할 것으로 생각된다.