1. 서 론
2. 연구 방법
2.1 조사 지역
2.2 데이터 수집
2.3 식물계절단계 추출
2.4 데이터 분석
3. 연구 결과
3.1 지소별 식물계절단계 관측 결과
3.2 연도별 식물계절 변화 경향 분석
3.3 기후요인 상관관계 분석
4. 고찰
4.1 식물계절 변화와 영향 요인
4.2 피노캠 관측 연구 방향 제안
1. 서 론
식물계절학 (Phenology)은 계절변화에 따른 식물의 생물학적 변화를 관찰하고 연구하는 학문이다 (Kramer et al. 2000). 주로 잎의 변색과 낙엽, 개화시기, 열매의 성숙 등과 같은 식물의 생물학적 사건들이 계절에 따라 어떻게 변화하는지를 기록하고, 이를 토대로 기후 변화에 따른 생태계의 반응을 이해하고 예측한다 (Piao et al. 2019). 이러한 식물계절현상은 환경 조건 변화에 민감하게 반응하며, 이러한 변화는 식물과 상호작용하는 다른 분류군에도 영향을 미쳐 전체 생태계 구조를 변화시킬 수 있다 (Duchenne et al. 2020, Gibson et al. 2016). 이 때문에, 기후변화가 심화됨에 따라 식물계절 변화에 대한 연구의 중요도 또한 높아지고 있으며 기후변화의 영향을 이해하기 위해 다양한 연구들이 수행되고 있다. Gordo and Sanz (2005)는 지중해 유역의 식물계절 변화가 기온 상승과 연관되어 있음을 밝혔고, Asse (2018)는 알프스 고산지대에서 겨울 및 봄 온난화가 식물의 개화 및 상호작용에 영향을 줄 수 있음을 확인했다. 또한 작물의 성장 시기 및 수확 시기를 예측하여 농업 생산성을 향상시키는데 활용하기도 하며 (Ren et al. 2019, Shim et al. 2010), 보전 및 복원 전략 수립을 위한 기초 자료 수립 등 (Kim et al. 2011) 다양한 분야에서 활발히 연구되고 있다.
가장 고전적인 식물계절 연구법은 직접적인 현장조사를 통해 식물계절현상을 기록하는 방법이다 (Aono and Kazui 2008). 이 방법은 현상을 정확하게 기록할 수 있다는 장점이 있지만 조사자의 접근이 힘든 극한지역에 적용하기 어렵고, 조사자 간의 주관이 개입할 여지가 크다는 단점이 있다 (Piao et al. 2019). 위성을 활용한 원격 탐사 기술의 발전으로 광범위한 영역에 대한 연구가 가능해졌으나, 기상 조건에 따라 데이터 품질이 크게 떨어질 수 있고 해상도가 낮아 개별 종 수준에서의 분석이 어렵다 (Tucker et al. 2010). 최근에는 이런 단점들을 보완하기 위해 RGB센서를 탑재한 자동영상촬영 카메라 (피노캠, Phenocam)를 활용하여 근거리에서 고빈도 (대략 30분 ~ 1시간 간격)로 이미지를 수집하여 식물계절 변화를 분석하는 연구방법이 급부상하고 있다 (Brown et al. 2016). 미국의 NEON (National Ecological Observatory Network), 호주의 TERN (Terrestrial Ecosystem Research Network) 등 해외 여러 장기생태연구기관에서 적극 활용하고 있으며, 한국의 국립생태원에서도 2017년 설치를 시작하여 현재(’23년 12월 기준) 총 8대의 피노캠을 운영하고 있다.
이처럼 식물계절연구를 위한 도구로써 피노캠의 중요도가 커졌음에도 불구하고 국내에서는 아직 관련 연구가 부족한 실정이다 (Han et al. 2020, Park et al. 2019). 본 연구에서는 국립생태원의 피노캠으로부터 수집된 이미지 데이터를 활용하여 국내 여러 산림의 식물계절변화 양상을 파악하고, 이에 대한 기후요인과의 상관관계를 규명하고자 하였다. 또한 피노캠을 활용한 연구법에 대한 개선점을 제안하여 기후변화에 대응하기 위한 식물계절자료 수립에 기여하고자 한다.
2. 연구 방법
2.1 조사 지역
국립생태원은 총 8개의 지소 (Table 1)에서 자동영상촬영장치 (Phenocam, 이하 피노캠)를 설치하여 식물계절변화 이미지 자료를 수집하고 있다. 본 연구에서는 점봉산, 서천, 우포, 완도, 한라산의 5개 지소에 설치된 피노캠 수집 자료를 분석하였다. 점봉산은 신갈나무군락 (Quercus mongolia) 및 소나무군락 (Pinus densiflora), 서천은 상수리나무군락 (Quercus acutissima) 및 곰솔군락 (Pinus thunbergii), 우포는 버드나무군락 (Salix koreensis), 완도는 붉가시나무군락 (Quercus acuta), 그리고 한라산은 구상나무군락 (Abies koreana)의 변화를 모니터링하고 있다 (Fig. 1, Table 1).
Table 1.
Information of Phenocam sites. GPS: global positioning system, AWS: the nearest Automatic Weather Station

2.2 데이터 수집
피노캠은 크게 이미지 데이터를 촬영하는 RGB센서 카메라 (Nikon d5600), 촬영스케줄, 무선통신 등을 자동으로 조정하는 컨트롤러, 데이터 원격전송을 위한 LTE 모뎀, 그리고 전원을 공급하는 태양열 패널로 구성되어있다 (Fig. 2). 모든 지소는 컨트롤러를 통해 07:00부터 19:00까지 한시간 간격으로 촬영되도록 설정되었으며 촬영된 이미지는 ftp 무선 전송 방식으로 국립생태원 데이터 PC로 수집되었다. 그림자에 의한 영향을 최소화하기 위해 해가 높게 떠 있는 11:30 ~ 13:30 사이에 촬영된 이미지 데이터를 선별하여 분석을 수행했으며, 각 지소의 연간 분석에 사용된 이미지 데이터는 Table 2에 정리하여 나타내었다.

Table 2.
Image data used in phenological analysis. It shows the annual count and total size of image data used for analysis in each phenocam site
2.3 식물계절단계 추출
수집된 이미지는 R 패키지 “Phenopix”를 이용하여 분석을 진행했다. Phenopix는 이미지 픽셀 기반으로 RGB DN (Digital Number, 디지털 넘버) 값을 추출하고 식생지수를 계산하는 패키지이다 (v2.4.2; Filippa et al. 2016a). 선별된 이미지 중 군락이 잘 구분되는 이미지를 대표 이미지로 선정하고 지소별 대표 군락이 포함되도록 관심구역 (Region of Interest, ROI)을 설정하였다 (Fig. 3). ROI 내의 모든 픽셀마다 RGB DN (디지털 넘버)값을 추출한 후, Gcc (Green Chromatic Coordinate) 값을 계산하여 식생 지수를 정량화하여 추출하였다 (Eq. 1). Gcc 계산식은 다음과 같으며, 매일의 ROI 내 전체 Gcc 평균값을 기록하여 연중 변화를 기록하였다.

Fig. 3
Region of Interest (ROI) defined for extracting RGB Digital Number for each sites. a) Wando, b) Upo, c) Jeombongsan Mt., d) Seocheon, e) Hallasan Mt.. Observation target is depicted in Table 1.
식물계절단계 추출에 앞서 안개, 비 등 기상 상황의 영향을 최소화하기 위해 패키지에서 제공하는 ‘night’, ‘spline’, ‘max’ 필터를 적용하여 Gcc 분포 그래프에서 이상치를 제거하였다. 또한 단일 관측값의 영향을 줄이고 계절적 변동을 더 잘 포착하기 위해 패키지에서 제공하는 이중 로지스틱 방정식 적합법 (Double logistic regression fitting method) 중 모든 지소 및 연도에 대해 평균적으로 가장 적은 평균 제곱근 오차 (RMSE, 평균 0.005)를 보이는 Klosterman 방법 (Klosterman et al. 2014)을 적용하였다. 식물계절단계 추출법은 연중 Gcc 변화가 뚜렷한 낙엽성 식물과 Gcc 변동이 작은 침엽수종 등을 아울러 적용할 수 있는 비교적 단순한 Trs 방법 (Threshold method)을 채택하였다. Trs 방법은 생장시작일: sos (start of season), 최대생장일: pop (peak of season position), 생장종료일: eos (end of season) 총 3단계의 식물계절단계를 추출한다 (Filippa et al. 2016b). pop는 Gcc가 최대에 도달하는 시기를 나타내며 sos와 eos는 각각 봄과 가을에 sos가 50%에 이르는 시기를 나타낸다. 위 과정은 연도별로 모든 지소에 대해 적용하여 식물계절현상의 변화를 측정하였다.
2.4 데이터 분석
각 지소의 연간 식물계절 변동이 유의한 경향성을 나타내는지 확인하기 위해 각 단계에 대해 선형회귀분석을 수행했다. 또한, 변동 경향과 기후요인이 상관관계를 가지는지 확인하기 위해 피어슨 상관 계수를 구해 측정하였다. 기상 데이터는 기상청 기상자료개방포털에서 제공하는 자동기상관측장비 (AWS)에서 2019년 1월 ~ 2023년 10월 동안의 월 평균기온, 월 최고기온, 월 최저기온, 월 누적강수량을 활용하여 분석하였다. 각 지소별로 가장 가까운 AWS에서 관측된 기상 데이터를 사용했으며 위치는 Table 1에 나타내었다. 기온은 조사지와 AWS의 고도차를 고려하여 (-0.6°C/100 m) 보정한 후 분석하였다. 모든 분석은 R version 4.2.3으로 수행되었다 (R Core Team 2023).
3. 연구 결과
3.1 지소별 식물계절단계 관측 결과
각 지소의 연중 Gcc 변화를 klosterman 방법으로 적합한 후 trs 방법으로 식물계졀현상을 추출한 결과를 나타내었다 (Fig. 4). 추출된 결과에서 생장종료일 (eos)에 생장시작일 (sos)를 뺀 값을 전체 생장 기간으로 정의하고 각 지소에 대해 연도별로 결과를 나타내었다 (Table 3). Jb-Qm의 경우 2022년 ~ 2023년의 성장시작일이 제대로 추정되지 않았으며, 서천 지소의 경우 통신상의 장애로 여름 기간의 데이터가 다소 누락되어 전체적으로 적절한 추정이 이루어지지 않았다. 한라산 지소는 2023년의 경우 악천후 조건에 의해 데이터가 충분히 수집되지 않아 분석에서 제외되었다.

Fig. 4
The analysis of phenophases by year for each site. The blue line represents sos, the green line represents pop, and the red line represents eos. Hl-Ak: Hallasan-Abies koreana, Jb-Qm: Jeombongsan-Quercus monglica, Jb-Pd: Jeombongsan-Pinus densiflora, Sc-Pt: Seocheon-Pinus thunbergii, Sc-Qa1: Seocheon-Quercus acutissima, Wd-Qa2: Wando-Quercus acuta, Up-Sk: Upo-Salix koreenis. *sos: start of season, pop: peak of season position, eos: end of season.
Table 3.
Yearly length of growing season in each phenocam site. It represents the value of end of season (eos) minus start of season (sos). Hl-Ak: Hallasan-Abies koreana, Jb-Qm: Jeombongsan-Quercus monglica, Jb-Pd: Jeombongsan-Pinus densiflora, Sc-Pt: Seocheon-Pinus thunbergii, Sc-Qa1: Seocheon-Quercus acutissima, Wd-Qa2: Wando-Quercus acuta, Up-Sk: Upo-Salix koreenis
2019 ~ 2023년 식물계절 결과값의 평균을 비교했을 때, 참나무과 낙엽활엽수림에 속하는 Jb-Qm과 Sc-Qa1은 생장시작일이 각각 130.0 ± 1.5, 120.0 ± 2.0, 최대생장일은 144.4 ± 1.2, 137.4 ± 2.1로 서 Sc-Qa1이 생장시작일은 약 10일, 최대생장일은 약 7일 가량 빨랐다. 생장종료일의 경우 각각 270.2 ± 6.7, 303.3 ± 3.5로 Jb-Qm가 약 33일 가량 더 빠르게 잎의 변색이 시작되는 것으로 나타나 전체적인 생장 기간은 Sc-Qa1이 한달 이상 길어 크게 차이가 나는 것으로 확인됐다. 소나무속 식물인 Jb-Pd와 Sc-Pt의 경우 생장시작일은 각각 118.8 ± 1.7, 144.5 ± 15.7로 Jb-Pd가 약 27일 가량 빨라 큰 차이를 보였고, 생장종료일은 282.2 ± 4.6, 320.8 ± 7.5로 Jb-Pd가 약 39일 빠른 것으로 나타났다. 전체 생장 기간은 Sc-Pt이 약 13일 길었고 Jb-Pd가 일찍 생장을 시작하지만 더 빨리 생장이 종료되는 것으로 확인됐다. Up-Sk의 경우 다른 지소에 비해 생장시작일이 약 80.8 ± 1.7로 가장 빨랐다. 생장종료일은 약 277.0 ± 4.3으로 전체 생장 기간은 약 196.3일로 다른 6개 지소에 비해 가장 길었다. 상록활엽교목에 속하는 Wd-Qa2는 생장시작일이 128.0 ± 5.5, 생장종료일이 242.4 ± 11.0 이며, 전체 생장 기간은 114.4일로 모든 지소 중 가장 짧았다. Hl-Ak는 생장시작일이 116.5 ± 29.6으로 변동폭이 매우 크게 나타났다. 생장종료일은 307.8 ± 6.4로 전체 생장 기간은 약 191.3일로 나타나 같은 침엽수종인 소나무속 식물에 비해 길게 나타났다. 전반적으로 같은 속 식물과 비교했을 때 위도가 높을수록 생장종료일이 빠르고 생장 기간이 짧은 경향이 있었다.
3.2 연도별 식물계절 변화 경향 분석
2019 ~ 2023년 동안 지소별 식물계절 변화의 경향성을 확인하기 위해 선형회귀분석을 수행한 결과를 나타내었다 (Fig. 5). 생장시작일은 Up-Sk (0.5 ± 2.0)와 결측치가 많아 분석이 어려운 Sc-Qa1을 제외하고 음의 회귀계수를 보여 해가 지남에 따라 점차 빨라지는 경향을 보였다. Hl-Ak가 -22.4 ± 12.5로 오차가 컸으나 가장 변화량이 크게 나타났으며, Sc-Pt가 -8.0 ± 12.0으로 두번째로 컸다. 최대생장일 또한 생장시작일과 비슷한 경향을 보였으나 그 변동은 적은 편이었다. 생장종료일의 경우 Hl-Ak (8.1 ± 4.1)와 Jb-Pd (1.9 ± 3.6)는 점차 늦어지는 경향을 보였으나 다른 수종은 모두 빨라지는 경향을 보였다. Jb-Qm가 -6.9 ± 3.7로 감소하는 정도가 가장 컸고, Up-Sk가 -6.2 ± 1.6으로 두 번째였다.

3.3 기후요인 상관관계 분석
지소별 식물계절의 변화와 기후요인과의 상관관계를 확인하기 위해 피어슨 상관분석을 수행하였다 (Fig. 6). Sc-Pt, Sc-Qa1 그리고 Jb-Qm은 결측치로 인해 표본 수가 부족하여 분석에서 제외하였다. Wd-Qa2의 경우 4월 최고기온과 생장시작일이 음의 상관관계를 보이고, 8월 최저기온이 생장종료일과 음의 상관관계를 보였다. Up-Sk는 겨울철 기온이 최대생장일과 상관관계가 있었으며, 1월 최고기온과 2월 최저기온이 낮을수록 최대생장일이 늦어지는 것으로 나타났다. 반면 Hl-Ak는 8월 평균기온과 최대생장일이 양의 상관관계를 보였고, 8월 강수량이 많을수록 생장종료일이 빨라졌다. Jb-Pd는 가장 많은 요인과 상관관계를 보였는데, 생장시작일은 1월 최고기온, 최대생장일은 1월 평균기온과 2월 누적강수량, 생장종료일은 7월 최고기온과 양의 상관관계가 있는 것으로 나타났다.

4. 고찰
4.1 식물계절 변화와 영향 요인
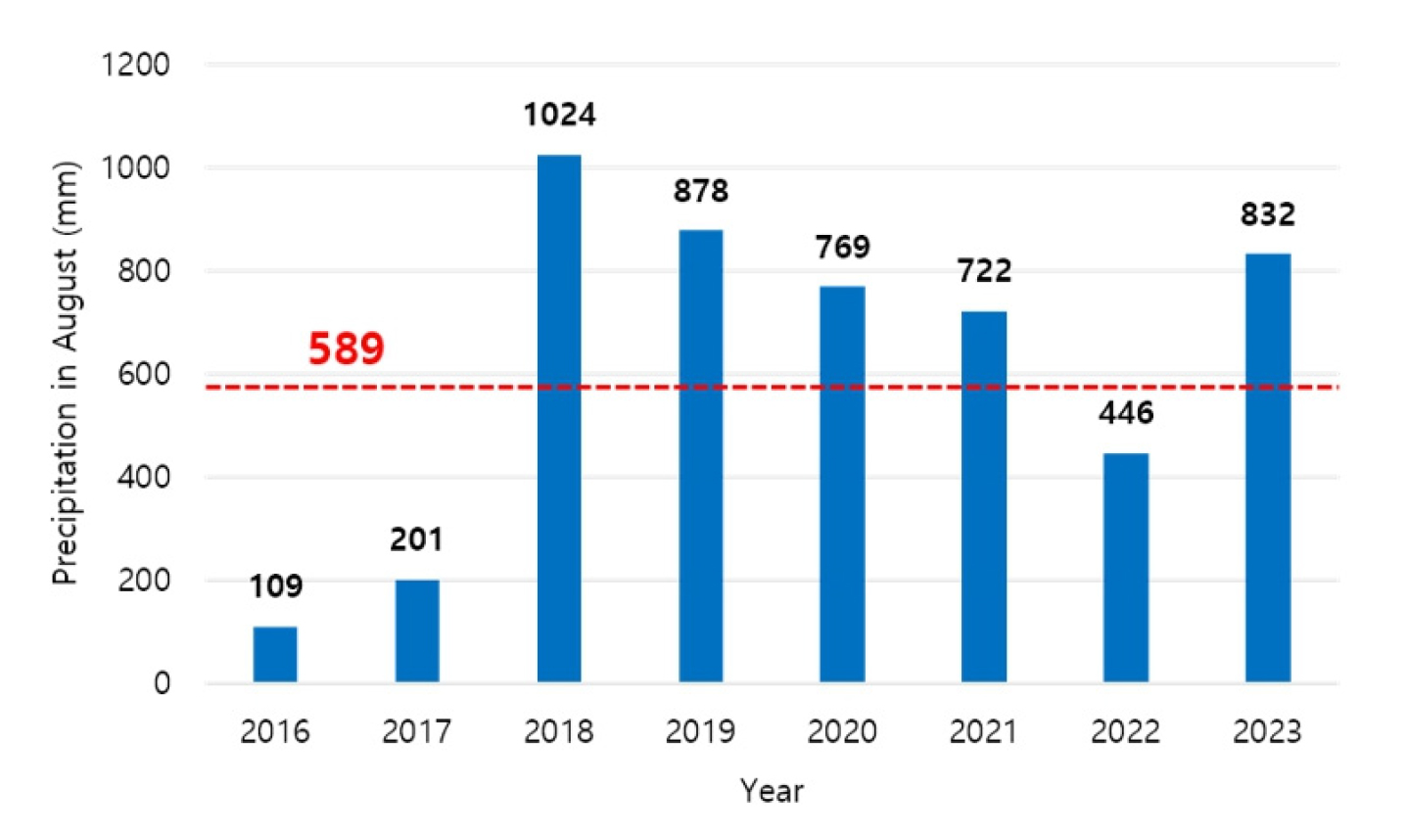
본 연구 결과에서 침엽수림들은 전반적으로 전체 생장 기간이 증가하는 경향을 보였고, 우포의 버드나무는 생장시작일이 늦어지고 생장종료일이 빨라지며 전체 생장 기간이 짧아지는 경향을 보였다. 그러나 변화되는 정도는 지소별로 상이하게 나타났다. 일반적으로 기후변화로 인한 기온상승은 잎의 노화를 지연시키는 것으로 알려져 있으나, 식물계절변화에는 더 복잡한 요인들이 관여하며, 종에 따라 그 정도가 다양하게 나타날 수 있다 (Estiarte and Penuelas 2015). 점봉산의 소나무의 경우 겨울철 기온 및 강수량이 생장 시작과 최대 생장 도달에 영향을 크게 미치는 것으로 나타나는데 이는 이른 봄에 녹는 눈이 수분 가용성을 높여 생장을 촉진하기 때문인 것으로 추측된다 (Schreiber et al. 2013, O’Leary et al. 2018). 한라산의 구상나무의 경우 8월 강수량이 많을수록 생장종료일이 빨라지는 것으로 나타났는데, 이는 최근 보고되는 구상나무 군락의 쇠퇴 현상과 관련이 있을 것으로 보인다 (Song et al. 2019). Ahn (2019)은 구상나무의 고사 현상이 토양 수분스트레스에 의한 것일 수 있음을 제안하였다. 국내 여름철 강수량은 기후변화에 의해 점차 증가하는 추세를 보이고 있는데 (Oh et al. 2022), 실제 한라산 영실 AWS에서 관측된 8월 강수량합은 2017년 200.5 mm에서 2018년 1024.0 mm로 약 5배 증가하였으며 2022년을 제외하고 2000년 ~ 2023년 측정된 한라산의 8월 강수량합 평균인 589 mm를 초과하였다 (Appendix 1). 본 연구 결과에서 8월 강수량합이 446 mm로 낮았던 2022년은 생장종료일이 327일로 2019년 ~ 2021년의 생장종료일보다 약 25일 늦게 나타나 구상나무의 생육에 영향을 줄 수 있는 것으로 판단된다. 따라서 구상나무 고사 현상의 원인을 파악하기 위해 여름철 강수량과의 관계에 대한 추가적인 연구가 필요할 것으로 보인다.
지리적 조건에 따라서도 식물계절 변화 양상은 달라질 수 있는데, 북미 지역에서 수행된 식물계절 연구에서 위도별로 기후변화에 대한 향후 시나리오 양상이 다르게 예측되었다 (Seyednasrollah et al. 2020). 영국의 낙엽수림에서 수행된 연구에 따르면 적당한 열 스트레스는 휴면 시기를 늦추는 결과를 보였으나, 추위 및 고온 스트레스는 오히려 휴면 시기를 앞당기는 모습을 보였다 (Xie et al. 2015). 본 연구에서는 지소별로 수종이 달라 정확한 비교는 할 수 없었으나 같은 수종이라도 환경 조건에 따라 식물계절 변화 양상이 달라질 수 있음을 나타내고 있다.
4.2 피노캠 관측 연구 방향 제안
식물계절 단계 추출 결과, 정확도 높은 분석을 위해선 결측치를 최소화하는 것이 중요하다. 서천 지소의 경우 전원 이상, 통신 장애 등 기계적인 오류 발생으로 여름철 결측치가 많이 발생하여 식물계절 단계 추출과 기후요인 상관관계 분석이 불가능 했다 (Fig. 4). 장시간 전원 공급이 차단되는 것을 방지하기 위해 가능한 전력선을 통한 전원과 태양열 전원을 동시에 사용하는 것이 권장되며 태풍, 폭우 등 극한 기상 현상이 있을 시 이후 즉각적인 점검이 필요하다. 2023년 한라산의 경우, 여름철 폭우와 겨울철 서리 등에 의해 분석에 사용할 수 있는 이미지 데이터의 양이 부족해 분석에 실패한 바있으므로 (Table 2, Fig. 4), 촬영 간격을 1시간에서 30분으로 단축하여 분석 해상도를 높이는 방안을 계획하고 있다. 또한 야외 환경에 노출되는 장비인 만큼 모뎀 장비 고장이 있을 수 있어 통신 장애에 대비하여 원격 전송이 실패했을 때 최소 2주 분량 정도의 데이터를 저장할 수 있는 예비 저장 장치를 현장에 연결해야 한다.
올해 새로 설치된 3개의 피노캠 (고양 장항습지, 서울 남산, 영양 멸종위기종복원센터)을 제외하면 현재 5년 이상 국립생태원에서 데이터를 수집한 지소는 5개에 불과하다 (National Institute of Ecology 2022). 단순한 현상 관찰에서 벗어나 지리적 요인, 기후적 요인 등을 고려한 고도화된 분석을 위해서 다양한 생태계 유형을 반영할 수 있도록 표준화된 관찰 지소를 지속적으로 확대할 예정이다. 동일 수종에 대해 위도, 고도, 서식지 등을 달리하여 추가 관찰 지소를 설치하여 지리적 요인에 대한 식물계절변화 양상의 차이를 분석할 수 있다. 예를 들어 한라산 구상나무의 식물계절변화 양상과 비교하기 위해 지리산 구상나무군락을 대상으로 관찰 지소를 추가한다면 8월 강수량의 영향이 다른 지역에서도 일반적으로 나타나는 현상인지 확인할 수 있다. 식물계절현상은 기후요인에 영향을 크게 받는 만큼 피노캠 설치 장소에 기상모니터링을 위한 장비를 같이 운용하는 것이 필요하다. 현재 국립생태원에서 설치한 피노캠 관찰 지소에는 대부분 기상모니터링 장비가 같이 설치되어 있어 이를 활용하여 보다 정확한 기후요인 분석을 진행할 예정이다. 추가 설치시에도 기온 및 강수량을 측정할 수 있는 장비를 병행하는 것이 필요하다.
최소 10년 이상 장기간 축적된 피노캠 관측 데이터는 생태계 기후변화의 기후변화 영향을 판단할 수 있는 중요한 자료로 활용될 수 있으며 선진국에서는 그 가치를 인정하고 다량의 데이터를 축적하고 있다. 또한 가시광선 영역 뿐만 아니라 빛의 파장대를 확대하면 좀 더 세밀한 식물의 생리적 변화를 판단하는 지표가 될 수 있을 것이다. 피노캠을 통해 얻어진 고해상도의 Gcc 기반 식물계절 데이터를 위성 데이터와 연계하면 생물계절 연구결과를 국가 규모로 확대하여 분석할 수 있다. (Burke and Rundquist 2021, Cui et al. 2019, Thapa et al. 2021). 피노캠의 Gcc는 위성 기반의 NDVI, EVI 등 다른 식생지수와 상관관계를 가지는 것으로 나타났고, 이를 바탕으로 분석 범위를 확장할 수 있을 것으로 보인다 (Khare et al. 2022). 전국 범위의 기후변화 대응 식물계절 기초 자료를 수립하기 위해 국내 생태계 유형을 대표할 수 있는 피노캠 관찰지소를 추가로 설치하고, 이를 위성 데이터와 결합하려는 연구가 지속되어야 한다.