1. 서 론
2. 연구 방법
2.1 조사지 개황
2.2 연구방법
2.3 자료 처리 및 분석
3. 결 과
3.1 식생군락의 분포 특성
3.2 환경요인과 주요식생 군락의 상관관계
4. 고 찰
4.1 토양 환경요인의 변화
4.2 식생 군집의 변화
4.3 기후변화에 의한 식생 영향
5. 결 론
1. 서 론
갯벌이란 조석 현상에 의해 주기적으로 침수를 겪는 만조선과 간조선 사이의 연안습지로서, 조차가 크고 경사가 완만하며, 파랑에너지가 낮은 것이 특징이다. 퇴적물의 입자 크기에 따라 모래 갯벌, 혼합 갯벌, 펄 갯벌로 구분되며, 바닷물에 의한 주기적 침수에 의해 염분과 침수에 대한 스트레스가 매우 높은 환경이다 (Fan et al. 2013, Richards et al. 2005). 이러한 스트레스 환경에 적응한 동식물 중에는 멸종위기 야생생물 2급인 흰발농게, 해양수산 생명자원인 두꺼비갯민달팽이 등의 서식이 연구되었다 (Kil and Lee 2011, Yoo et al. 2021). 또한, 대표적인 국내 염생식물인 칠면초, 갈대 등이 갯벌 서식지에 미치는 영향에 대한 연구들이 진행되어 왔다 (Bang et al. 2020, Kwon and Je 2002, Shin et al. 2020). 이 외에도 멸종위기 야생생물 2급인 검은머리물떼새, 알락꼬리마도요, 1급종인 저어새 등의 철새가 갯벌의 저서무척추동물이나 염생식물의 근경 및 괴경 등을 먹이로 삼아 머물거나 월동하기도 한다 (Choi et al. 2017). 이러한 생태적 가치로 인하여 전라남도 순천과 신안, 전라북도 고창 그리고 충청남도 서천까지 연결되는 서해안의 갯벌은 2021년 세계자연유산으로 지정되었다 (Jeong 2021). 함평만은 무안-신안과 연결되어 서해안 갯벌을 구성하고 있다. 최근 신안군은 ‘갯벌 세계자연유산 보전본부’를 유치하였으며, 세계유산위원회는 기선정된 4곳 외에 2025년까지 갯벌 유산구역 확대를 권고하고 있어, 지리적으로 가까운 함평만 갯벌의 세계유산지정은 지자체의 노력과 함께 곧 실현될 가능성이 높다 (Kim 2022).
함평만은 함평군에서 무안군 및 영광군까지 이어지는 약 1만 ha에 이르는 갯벌에 위치하고 있다. 서남해안의 리아스식 해안을 대표하는 반폐쇄적인 만으로, 좁은 만 입구와 넓은 조간대 분포로 독특한 지형 구조로 인하여 비대칭적인 퇴적물 이동이 일어나는 곳이다 (Lee et al. 1998, Lee and Jun 2009). 함평만 연안 수심은 5 m 이하로 낮고, 평균 조위가 4.4 m로 간조 시 드러나는 넓은 갯벌은 모래 혹은 모래와 점토가 혼합된 저질로 조간대 하부로 갈수록 점토의 함량이 증가하는 것으로 연구되었다 (Park et al. 2017).
기후변화로 인한 해수면의 변동은 그 자체로 갯벌에 직접적인 영향을 줄 뿐 아니라 (Ministry of Environment 2020), 갯벌로 공급되는 퇴적물의 이화학적 성질을 변화시켜 염습지 생태계의 동식물상 및 규조류 등 저서생물상 변화에 영향을 미친다 (Gedan et al. 2009, Lee and Jung 2011, Lee 2013, So et al. 2019). 특히 염생식물은 그늘 제공, 생물학적 배수 기능, 염분 스트레스 완화 등을 통해 환경을 개선하여 서식지를 형성하는 역할을 하기 때문에 보전 전략을 수립함에 있어서 최우선적으로 고려해야할 대상이다 (Shamsutdinov et al. 2017). 따라서, 본 연구는 복잡한 해안지형으로 비대칭적 퇴적물 이동이 일어나는 함평만에서 염습지 식생군락과 토양 환경요인의 변화를 파악하고, 식생과 토양 환경요인 사이의 상관관계를 규명하고자 하였다.
2. 연구 방법
2.1 조사지 개황
함평만은 우리나라 서해 남부에 위치한 반폐쇄성 내만으로 총 길이는 약 17 km, 최대 폭은 12 km이다. 연구지역은 함평만의 가장 안쪽에 위치한 전라남도 무안군 현경면 현화리 (N 35°02’07.53”, E 126°25’07.53”)이다 (Fig. 1). 연구지역 일대는 2001년 국토해양부 습지보호구역으로 지정되었으며, 2008년에 ‘람사르습지’ 목록에 등재되었다 (Yook 2015, Lee et al. 2016). 독특한 해안구조로 인해 다양한 지형과 염생식물 군락이 잘 발달된 함평만은 ‘국가 장기생태연구’를 통하여 식생변화가 모니터링되었다 (National institute of Ecology 2016 and 2021). 특히 현화리 일대 갯벌은 함평만 내 염생식물의 분포가 넓은 지역 중 하나로서, 사구식물과 염생식물의 군락 분포가 뚜렷하게 구분되는 것이 특징적인 지역이다. 기후요인이 미치는 영향을 파악하기 위해 함평만 갯벌과 가장 가까운 기상관측지점인 목포관측소의 2016년부터 2022년 기온 및 강수량 자료를 확인하였다. 기상자료는 부록으로 첨부하였다 (Appendix 1).
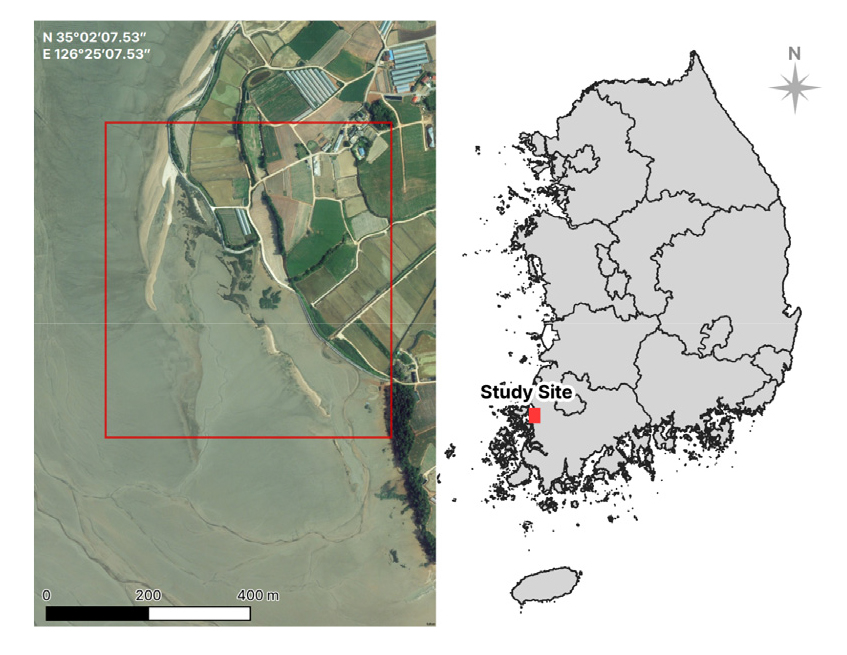
Fig. 1
The location of study site in Hampyeong Bay, Hyeongyeong-myeon, Hampyeong-gun, Jeollanam-do, Republic of Korea.
2.2 연구방법
2.2.1 현존식생도와 식생단면도 조사
현존식생도와 식생단면도를 작성하기 위해 2022년 7-10월에 현장조사를 수행하였다. 현존식생도는 함평만에 위치한 현화리 일대 염생식물과 사구식생이 발달된 지역을 중심으로 조사하였다. 식생도의 군집별 면적은 QGIS ver. 3.22 (QGIS.org 2022)을 이용하여 산출하였으며, 간조 시에 조사하여 갯벌에 발달된 전체 식생이 모두 포함되도록 하였다.
식생단면도 조사는 조사지 내 지형과 주요 식생인 갈대 (Phragmites australis), 갯잔디 (Zoysia sinica), 해홍나물 (Suaeda maritima) 등이 반영되도록 총 3개의 장소를 선정하여 띠조사구 방법 (transect)으로 조사하였다. 모든 띠조사구는 제방 바로 아래에서 시작하였으며 (Fig. 2 (a)), A-A' 조사구는 해홍나물 군집, B-B'는 갯잔디 군집, C-C'는 갈대 군집을 지나도록 설정하였다. 조사는 식생 피도가 5% 이하로 출현율이 낮은 지점에서 종료하였으며, 띠 조사구 A, B, C의 길이는 각각 119 m, 147 m, 그리고 175 m로 조사되었다 (Fig. 3 (2022)). 또한 조사지역의 지형을 시각화하기 위해 고정밀 지형 측량기 (TOPCON AT-B3)를 사용하여 시작점 높이를 기준으로 1 m 간격으로 지형의 상대고도를 측정하였다. 그리고 식생이 분포하기 시작하는 점과 끝나는 지점을 기록하여 거리별 식생 분포와 지형정보를 식생단면도로 나타내었다.

Fig. 2
Pictures of the field survey at Hampyeong Bay in 2022. (a) transect method (b) plot sampling (1 m × 1 m size).
2.2.2 군집조사
함평만 내 출현하는 우점종은 해홍나물, 갯잔디 및 갈대군락으로 파악되었다. 각 식생이 잘 발달된 지점에 1 m × 1 m 방형구를 3개씩 설치하여, 출현종과 피도 및 생물량을 조사하였다 (Fig. 2 (b)). 각 종의 식물체 높이는 최대 생장시기가 포함되는 시기에 조사하였다. 식물 동정은 원색 대한식물도감 (Lee 2003)을 기준으로 하였으며, 학명과 국명은 국립생물자원관의 ‘2021 국가생물종목록’을 따랐다 (National Institute of Biological Resources 2022). 생물량 (biomass) 조사는 방형구 내 우점종을 모두 수확하여 실험실에 운반한 후 토양 등 이물질을 제거하였다. 이후 80˚C 에서 3일 이상 건조하여 항량에 도달한 후 건중량을 측정하였다.
2.2.3 토양 환경요인 분석
토양 환경요인과 염생식물종의 상관관계를 파악하기 위해 주요 군집인 해홍나물과 갯잔디군락의 방형구마다 임의의 세 지점에서 표토를 채취하였다. 7월과 8월에 각각 채취하여 군락별로 총 6개의 샘플을 채집하였으며 채집 시기에 따른 차이는 없었다. 토양 환경요인으로는 수분함량 (moisture content, MC), 유기물함량 (organic matter, OM), 토성 (soil texture), 토양산도 (pH), 전기전도도 (electrical conductivity, EC), 총인 (total phosphorus, TP) 및 총질소 (total nitrogen, TN)를 측정하였다. 수분함량은 생토의 무게를 잰 후 도가니에 토양 샘플을 넣어 105˚C 에서 24시간 건조시켜 무게의 차이를 계산하여 백분율로 나타냈다. 유기물함량은 550˚C 작열기에서 4시간 동안 가열한 후 무게를 측정하여 작열손실량을 계산하였다. 토성은 입자의 크기가 0.05 mm-2.0 mm이면 모래 (sand), 0.05 mm 이하는 ‘미사+점토 (silt and clay)’로 구분하였다 (Kroetsch and Wang 2008). 토양산도와 전기전도도 측정을 위해 음건토양을 2 mm 체에 친 후 7 g을 증류수와 1:5 (w/w)로 혼합하여 60분간 진탕하였다. 진탕액을 100 µm mash에 여과한 후 pH-EC meter (YSI 60 pH & temperature, YSI pro30)로 측정하였다. 총인과 총질소의 분석은 경기환경과학연구원에 의뢰하였다. 각각 자동증류장치 (Vapo 200)와 흡광광도계 (UV-1800)를 이용하여 과염소산 황산 분해법으로 분석하였다 (National Institute of Crop Science 2014).
2.3 자료 처리 및 분석
2016년부터 2021년의 자료는 국가 장기생태연구 자료를 활용하였다 (National institute of Ecology 2016 and 2021). 시간에 따른 군락 면적 변화를 분석하기 위해 선형회귀분석 (linear regression)을 수행하였다. 유의확률 0.05 이하이면, 통계적으로 유의한 것으로 판단했으며, 그래프에 회귀식과 결정계수 (R2)를 같이 표기하였다. 2022년에 조사한 식생 군집 내 종별 기여도를 분석하기 위하여 중요도 (Importance value)를 계산하였다. 중요도의 계산은 각 군락의 종별 상대피도와 상대빈도 및 상대밀도의 합으로 계산하는 것이 일반적이다 (Curtis and McIntosh 1951). 그러나 연구대상종인 염생식물의 밀도를 반복적으로 조사하기에 시간적 제약이 따르므로 상대피도와 상대밀도 두 가지 값으로만 중요치를 계산하였다 (Bray and Curtis 1957). 또한 각 종의 중요도를 비교하기 쉽도록 상대피도와 상대빈도를 합한 값을 2로 나누어 최대값이 100이 되도록 표현하였다. 2022년 식생 군집별 토양 환경요인의 차이를 분석하기 위해 항목별로 등분산성을 확인한 후 t-검정을 수행하였다. 검정 결과 유의확률을 수준에 따라 표에 나타내었다. 군집의 분포와 토양 환경요인의 관계를 분석하기 위해 주성분분석 (PCA, principal component analysis)을 수행하였다. 주성분분석 결과는 행렬도 (biplot)로 나타내었으며, 주성분에 각 변인이 기여하는 정도를 확인하기 위해 factoextra package를 활용하여 그래프로 나타냈다 (v1.0.7; Kassambara and Mundt 2020). 또한, 변인들간의 상관관계를 확인하기 위해 피어슨 상관계수 방법을 이용하여 상관계수를 구하고 표로 나타내었다. 모든 통계분석은 R ver.4.2.1을 이용하였다 (R Core Team 2022).
3. 결 과
3.1 식생군락의 분포 특성
2022년 조사 결과, 함평만 현화리 갈대군락은 제방을 따라 약 100 m 길이에 최대 30 m 폭의 대상분포가 확인되었다 (Fig. 3). 갈대군락의 평균 높이는 2.5 m 정도였으며, 피도 100%의 순군락을 이루고 있었다 (Table 1). 식생단면도에서 확인할 수 있는 것처럼 갯벌에서 지형이 가장 높은 곳에 위치하고 있었으며, 갯잔디군락보다 약 10 cm, 해홍나물 보다는 약 30 cm 가량 지형이 높은 것으로 파악된다 (Fig. 4).
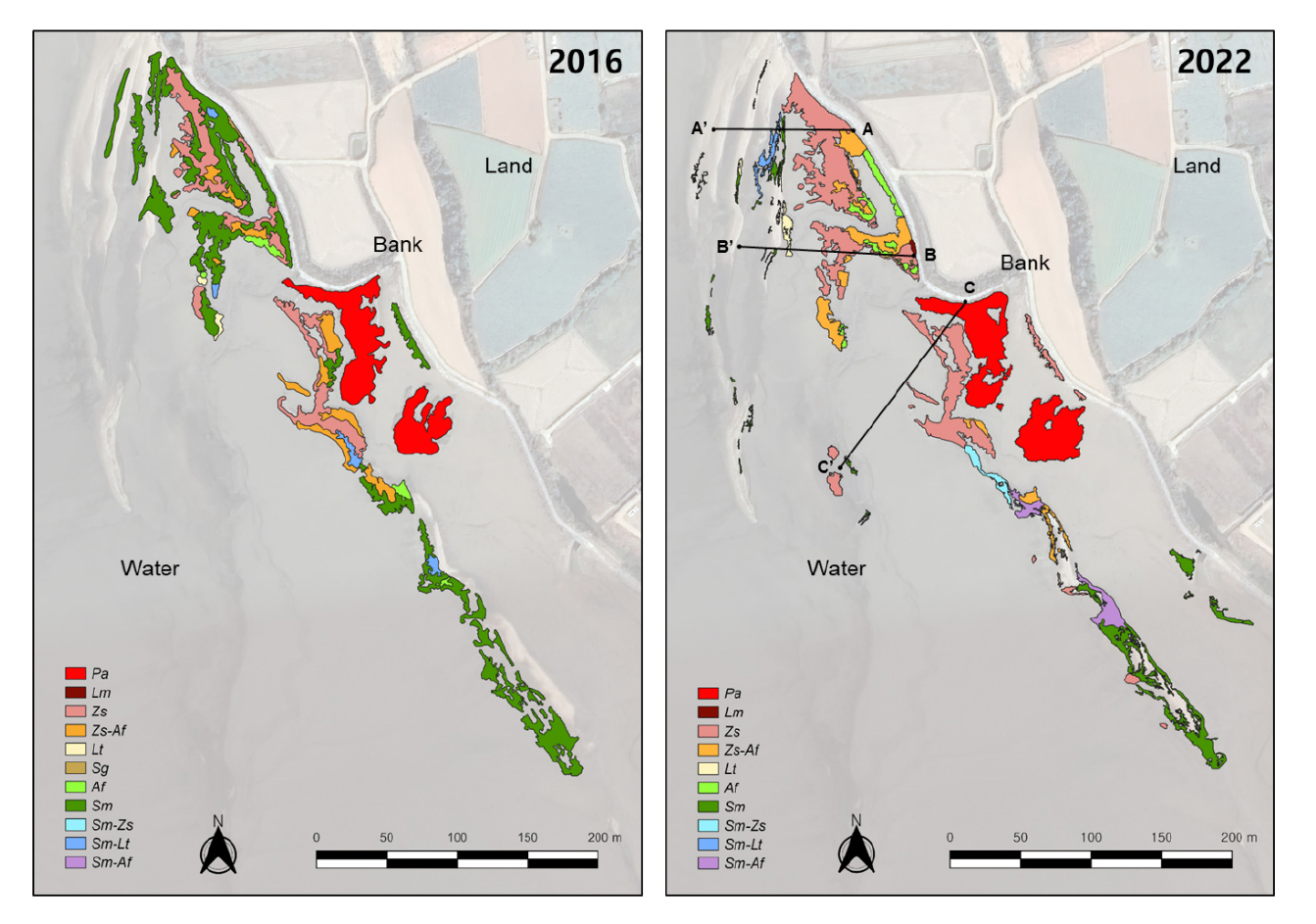
Fig. 3
Comparison of vegetation maps at Hampyeong Bay between 2016 and 2022. The transects indicated by 3 black lines installed in different vegetation types (A- A', B-B' and C-C'). Af: Artemisia fukudo, Lm: Leymus mollis, Lt: Limonium tetragonum, Pa: Phragmites australis, Sg: Suaeda glauca, Sm: Suaeda maritima, Zs: Zoysia sinica.
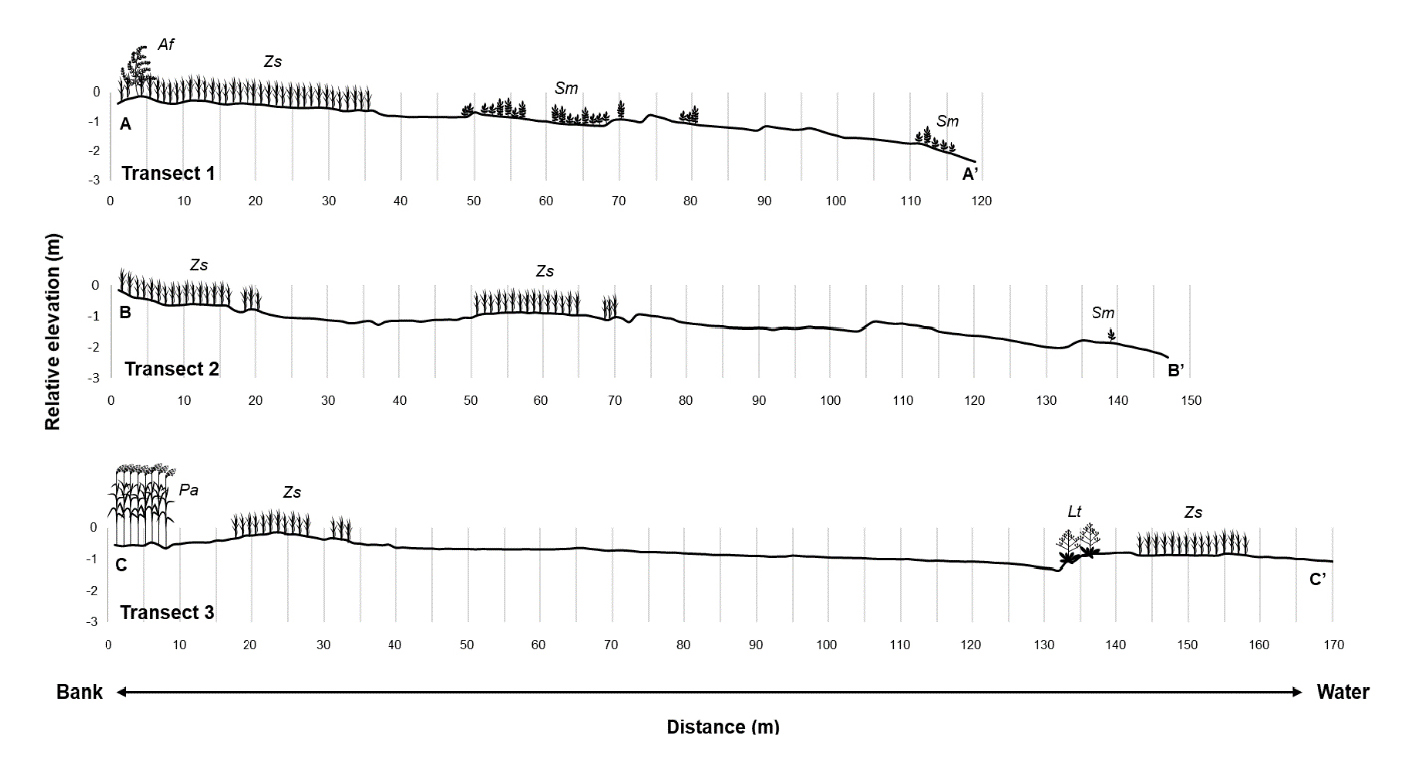
Fig. 4
Elevational profile of the three transects at Hampyeong Bay in 2022. Af: Artemisia fukudo, Lt: Limonium tetragonum, Pa: Phragmites australis, Sm: Suaeda maritima, Zs: Zoysia sinica.
Table 1.
Relative coverage, relative frequency and importance values by species within five representative vegetation types in 2022
|
Vegetation type
|
Species
|
Relative coverage (%)
|
Relative frequency (%)
|
Importance value (%)
|
| Limonium tetragonum (Lt)
| L. tetragonum |
72
|
43
|
57
|
| S. maritima |
25
|
43
|
34
|
| Z. sinica |
4
|
14
|
9
|
|
Total
|
100
|
100
|
100
|
Suaeda maritima
(Sm) | S. maritima |
100
|
100
|
100
|
|
Total
|
100
|
100
|
100
|
Zoysia sinica
(Zs) | Z. sinica |
93
|
75
|
84
|
| A. fukudo |
7
|
25
|
16
|
|
Total
|
100
|
100
|
100
|
Artemisia fukudo
(Af) | A. fukudo |
54
|
38
|
46
|
| Z. sinica |
40
|
38
|
39
|
| L. tetragonum |
5
|
13
|
9
|
| S. maritima |
1
|
13
|
7
|
|
Total
|
100
|
100
|
100
|
Phragmites australis
(Pa) | P. australis |
100
|
100
|
100
|
|
Total
|
100
|
100
|
100
|
갯잔디군락은 제방부터 해안까지의 구배 중 중간지점 이전의, 제방에 가까운 쪽에 주로 분포하는 것이 확인되었다 (Fig. 3). 갯잔디군락의 평균 높이는 22 cm 정도였으며, 피도 약 93%로 우점했다 (Table 1). 일부 지역에서는 큰비쑥 (Artemisia fukudo), 해홍나물 및 갯길경 (Limonium tetragonum)과 같은 다른 염생식물종들과 혼합군락을 형성하기도 했다. 갯잔디의 위치는 갈대보다 낮고 해홍나물보다 약 50 cm 가량 높은 것으로 파악된다 (Fig. 4).
해홍나물군락은 제방부터 해안까지의 구배 중 중간지점 이후의, 해안에 가까운 쪽에 주로 분포하였으며 일부는 제방을 따라 분포하는 것이 확인되었다 (Fig. 4). 해홍나물군락의 평균 높이는 약 11 cm 정도이며, 피도 100%의 순군락을 이루고 있었다 (Table 1). 해홍나물은 주로 고도가 상대적으로 낮은 해안쪽에서 출현했다 (Fig. 4).
2016년과 비교하여 2022년에는 해홍나물 순군락의 약 74%가 감소하고 갯잔디 순군락은 75% 증가하였다 (Table 2). 조사지역 중 북쪽의 해홍나물 대군락은 2022년에 갯잔디 대군락으로 대체되었다 (Fig. 3). 해홍나물군락들은 파편화되어 군락의 크기가 감소하거나, 다른 염생식물 종들과 혼합군락이 되어 전체적으로 면적이 시간에 따라 감소하는 양상이었으며 (Fig. 3 and 5) 이러한 경향이 통계적으로 유의했다 (p <0.01). 반면 갯잔디 순군락의 면적은 시간에 따라 증가하는 경향이 통계적으로 유의했고 (p <0.05), 갈대군락은 2016년부터 일정 면적을 유지하는 경향이었다 (Fig. 5).
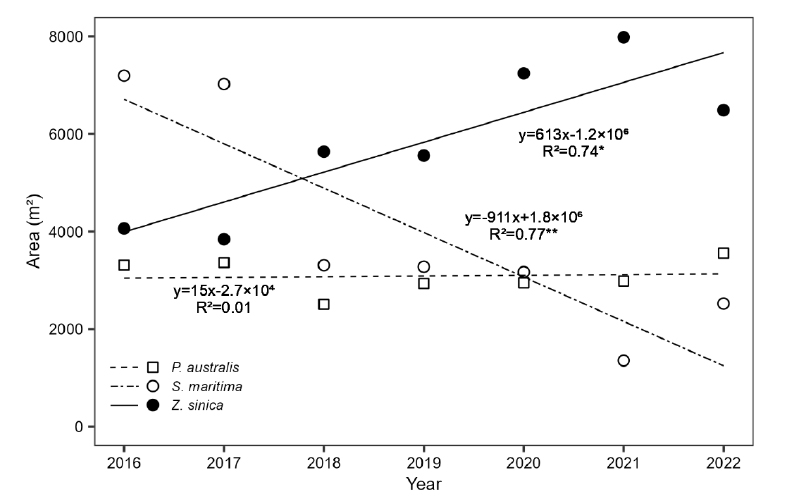
Fig. 5
Changes in vegetation area over time from 2016 to 2022 at Hampyeong Bay. *: p-value <0.05, **: p-value <0.01.
Table 2.
Comparison of community area in the existing vegetation map between 2016 and 2022
|
Vegetation type
|
Area in each year (m2)
|
Change (%)
|
|
2016
|
2022
|
| Pa |
3312
|
3556
|
7
|
| Lm |
0
|
25
|
-
|
| Zs |
3037
|
5302
|
75
|
| Zs-Af |
1464
|
1599
|
9
|
| Lt |
92
|
183
|
98
|
| Sg |
31
|
0
|
-100
|
| Af |
288
|
559
|
94
|
| Sm |
6979
|
1835
|
-74
|
| Sm-Zs |
0
|
226
|
-
|
| Sm-Lt |
311
|
208
|
-33
|
| Sm-Af |
0
|
547
|
-
|
3.2 환경요인과 주요식생 군락의 상관관계
2016년도와 2022년도 함평만 염생식물 군락의 토성을 비교하였다 (Fig. 6). 두 군락 모두 2022년에 모래의 비중이 크게 증가하였다. 두 군락 중 해홍나물군락의 모래 비율이 갯잔디군락보다 유의하게 높았다 (p <0.001, Table 3). 현장에서 관찰하였을 때 해홍나물군락은 대부분 사구화가 진행된 양상이었으며, 갯잔디군락의 사구화 진행속도는 해홍나물군락에 비해 더딘 것으로 나타났다.
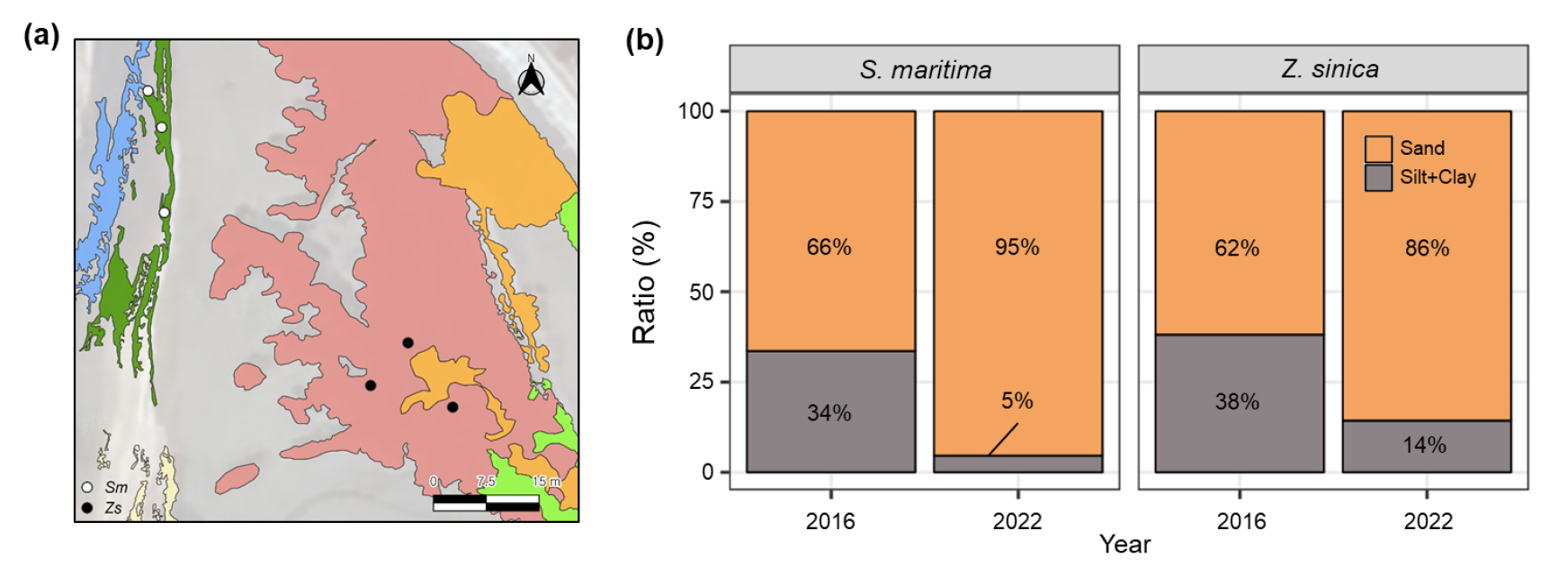
Fig. 6
(a) Soil sampling points. Sm: Suaeda maritima, Zs: Zoysia sinica. (b) Comparison of soil texture in S. maritima and Z. sinica between 2016 and 2022.
Table 3.
Comparison of soil variables between S. maritima stand and Z. sinica stand in 2022
|
Variable (Unit)
| S. maritima (n=6)
| Z. sinica (n=6)
| p-value
|
|
Mean
|
SE
|
Mean
|
SE
|
|
Moisture content (%)
|
3.8
|
0.4
|
12.3
|
0.7
|
***
|
|
Organic matter (%)
|
0.34
|
0.02
|
1.49
|
0.10
|
***
|
|
pH
|
7.56
|
0.04
|
6.99
|
0.02
|
***
|
|
Conductivity (mS/m)
|
445
|
78
|
1382
|
310
|
*
|
|
Sand ratio (%)
|
95.4
|
0.4
|
85.7
|
0.5
|
***
|
|
Total N (mg/L)
|
1050
|
150
|
950
|
193
|
n.s
|
|
Total P (mg/L)
|
58
|
7
|
127
|
14
|
**
|
2016년과 2022년 해홍나물군락과 갯잔디군락의 토양 환경요인의 차이를 t-검정으로 분석했다 (Table 3). 토양수분함량, 유기물함량, 전기전도도 및 총인의 값은 해홍나물군락보다 갯잔디군락에서 유의하게 더 컸다. 이와 달리 토양산도는 해홍나물군락에서 갯잔디군락보다 유의하게 높았는데, 이는 해홍나물군락이 해안 가까이 위치하여 해수의 영향을 받았기 때문인 것으로 판단된다.
함평만 내 염생식물 군집 분포에 영향을 미치는 토양 환경요인을 분석하기 위해 주성분분석 (PCA)을 실시하였다 (Fig. 7 (a)). 제1축에 따라 갈대-갯잔디-해홍나물 순서로 군집이 뚜렷하게 구분되었다. 수분함량, 유기물함량, 총인 및 전기전도도가 높은 곳에 갈대군락이 분포하는 것으로 분석되었다. 반면 앞에서 언급한 네 요인의 값이 낮고, 모래의 비율과 pH가 높은 곳에 해홍나물군락이 분포하는 것으로 분석되었다. 토양 환경요인들은 대부분 제1축의 분산에 비교적 고르게 기여하고 있었다 (Fig. 7). 한편 제2축의 분산에는 총질소의 기여도가 전체의 약 60%로 다른 요인들보다 큰 비중을 차지하고 있었다. 분석에 이용한 토양 환경요인간 상관관계를 분석하였다 (Table 4). 모래 비율은 수분함량, 유기물함량, 전기전도도, 총인과 강한 음의 상관관계를 나타내었다. 제2축 전체 분산의 대부분을 차지하는 총질소는 전기전도도와 음의 상관관계를 보였으나, 다른 변인과는 유의한 관계는 없는 것으로 나타났다.
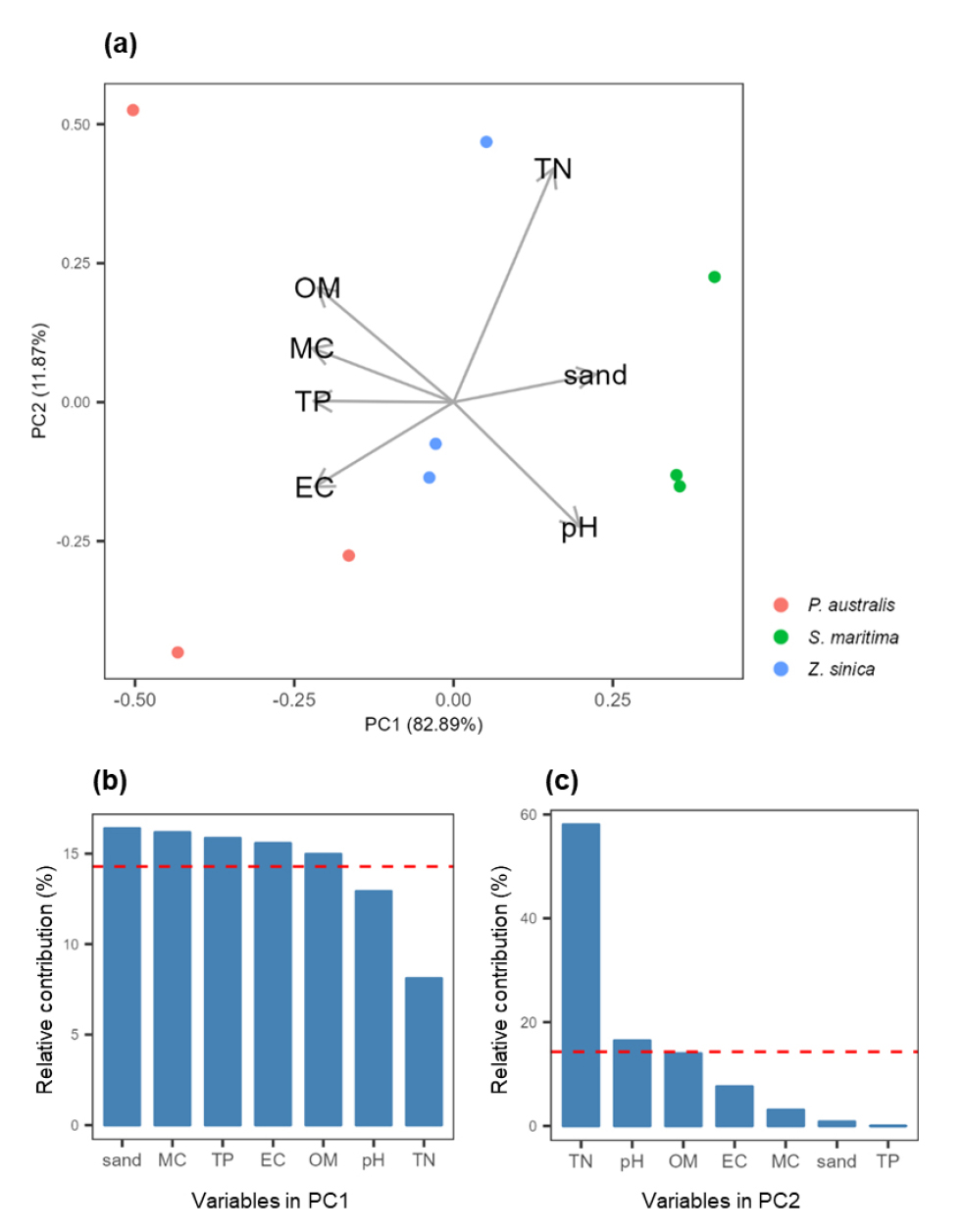
Fig. 7
Analysis of the relationship between soil variables and three halophyte species. (a) As a result of PCA, axis 1 variance was 82.89% and axis 2 variance was 11.87, explaining 94.76% of the total cumulative variance. The length of the arrow indicates the size of each variable's influence. (b) Relative contribution of factors influencing axis 1. (c) Relative contribution of factors influencing axis 2. The red dotted lines in (b) and (c) are the average values of variables affecting each axis. OM: organic matter, MC: moisture content, TP: Total Phosphorus, EC: Electrical conductivity, sand: ratio of sand, TN: Total Nitrogen.
Table 4.
Correlation between soil factors in 2022. Sand: ratio of sand, TN: Total Nitrogen, TP: Total Phosphorus, EC: Electrical conductivity, MC: moisture content, OM: organic matter
| Sand |
0.68*
|
-0.95***
|
0.77*
|
-0.97***
|
-0.93***
|
-0.87**
|
| TN |
-0.64
|
0.40
|
-0.80**
|
-0.54
|
-0.41
|
| | TP |
-0.79*
|
0.88**
|
0.91***
|
0.90***
|
| | | pH |
-0.71*
|
-0.86**
|
-0.92***
|
| | | | EC |
0.90***
|
0.79*
|
| | | | | MC |
0.97***
|
| | | | | | OM |
4. 고 찰
4.1 토양 환경요인의 변화
좁은 만 입구와 복잡한 지형으로 조성된 함평만은 지리적 여건으로 인하여 반폐쇄적인 구조를 하고 있다. 독특한 지리조건은 비대칭적인 퇴적물의 이동을 발생시키는 것으로 연구되었다 (Lee et al. 1998, Lee and Jun 2009). 퇴적물의 물리적 이동은 그 자체로 갯벌에 직접적인 영향을 주며, 이차적으로는 퇴적물과 유입되는 영양물질의 변화로 갯벌의 이화학적 성질에도 영향을 미쳐 염습지 생태계의 동식물상 및 규조류 등 저서생물상까지 달라지는 것으로 연구되었다 (Lee and Jung 2011, Lee 2013, So et al. 2019).
함평만 연안 수심은 5 m 이하로 낮고, 평균 조위가 4.4 m로 간조 시 드러나는 넓은 갯벌은 모래 혹은 모래와 점토가 혼합된 저질로 조간대 하부로 갈수록 점토의 함량이 증가하는 특성이 있다 (Park et al. 2017). 지형적으로 높낮이가 크지 않은 넓고 완만한 갯벌조건으로 인하여, 지속적인 모래의 유입은 함평만 전체의 토성 변화로 이어질 수 있다.
4.2 식생 군집의 변화
모래의 유입에 따른 갯벌 지형구조는 토양 환경요인을 변화시켰다. 지속된 퇴적으로 높아진 지형은 식물의 적합서식지 조건을 바꿔 기존 식생이 사라지거나 대체되는 결과로 나타났다. 2016년 31 m2로 조사된 퉁퉁마디 (Salicornia europaea)는 절대염생식물 중 염분내성이 가장 높은 종이며, 함평만의 조간대에서 제일 하위에 위치하였으나 점차 사라져 2022년 조사에서는 확인되지 않았다 (Appendix 2). 또 다른 절대염생식물인 해홍나물은 그 면적이 절반 수준으로 감소하였으며, 감소된 면적은 대부분 갯잔디로 대체되었다. 본 연구에서 함평만 갯벌 전체적으로 모래 함량이 증가된 것을 확인할 수 있었는데, 모래화된 갯벌이 갯잔디에게 유리한 조건인 반면, 해홍나물에게는 불리하게 작용한 것으로 판단된다.
특이할 만한 상황은 갯잔디가 모래화 조건을 더 발달시킨다는 점이다. 지하경이 발달되고, 지상부의 줄기가 복잡하게 얽혀 군락의 밀도가 높은 갯잔디는 모래와 같은 퇴적물의 이동을 방해하여 군락 내에 퇴적을 가속화시켰다 (Ganthy et al. 2013). 모래의 유입으로 사구가 유지되고, 갯잔디는 사구에 모래를 빠져나가 못하게 막아서 사구환경을 더욱 견고하게 지지하는 양상이다. 이러한 과정이 반복되어 모래가 지속적으로 퇴적되면, 향후 갯잔디가 있는 곳의 지형 고도가 증가할 것으로 예상된다. 이에 따라 점점 조간대 상부 지역은 침수 스트레스가 낮아지는 방향으로 바뀔 수 있다. 갯잔디는 상대적으로 침수 스트레스가 낮은 조간대 상부에 주로 서식하는 것으로 알려져 있다 (Bang 2018). 모래의 유입과 갯잔디군락의 상호작용은 갯벌 생태계의 변화를 파악하고 예측하기 위해 지속적으로 모니터링되어야 한다.
4.3 기후변화에 의한 식생 영향
기후변화는 평균기온 상승, 강우 패턴 변화, 가뭄이나 태풍 같은 기후적 현상의 빈도 변화를 동반하는데, 이는 염생식물의 군집 분포와 번식에 영향을 줄 수 있다 (Bernhardt and Leslie 2013, Etterson and Mazer 2016). 한국 기후변화 평가보고서 (Ministry of Environment 2020)에 따르면, 우리나라 주변 해역은 최근 49년간 (1968년 - 2016년) 표층 수온이 약 1.23°C 상승하였다. 이러한 온도의 상승은 목포관측소의 2016년부터 2022년의 평균기온, 최저기온, 최고기온이 모두 완만하게 상승하는 경향으로도 기후변화를 확인할 수 있다 (Park et al. 2022, Appendix 1). 기후변화의 영향은 온도 상승 이외에도 강수량에도 변화를 가져왔다. 우리나라는 2017년에 심한 가뭄을 겪었다 (Korea National Climate Data Center 2022, Appendix 1). 2016년부터 2022년까지 평균 총 강수량은 약 1,140 mm였으나, 2017년에는 약 723 mm로 최근 7년 중 가장 낮은 강수량을 기록했다. 특히 봄 철 강수량은 2017년에 89 mm로 같은 기간 연평균 256 mm의 1/3 정도에 불과한 것으로 조사되었다. 이렇게 가뭄으로 인해 토양 수분함량은 감소하고 토양 내 염분도가 증가하면 식물의 종자 발아에 악영향을 미칠 수 있다 (Anderson et al. 2022). 종자 번식에 의존하는 해홍나물이 2017년 이후 급격하게 그 면적이 감소한 것은 2017년 봄철 가뭄스트레스를 겪었기 때문일 수 있다. 함평만 인근의 목포관측소에서 2022년 11월까지 관측된 총 강수량은 약 717 mm이며, 봄철 평균 기온은 30년 평년 (1991-2020년) 값보다 1.5 ˚C 높은 것으로 조사되었다. 이러한 환경은 2017년과 비슷한 수준의 가뭄 조건을 형성할 수 있으며, 향후 함평만 해홍나물군락의 면적 감소 현상이 지속될 것으로 예상할 수 있다.
5. 결 론
함평만의 복잡한 지형 조건은 비대칭적 퇴적을 발생시켜, 지속적으로 퇴적물의 조립질화가 진행되고 있는 것으로 연구되었다. 갯벌의 모래 퇴적물은 지형을 높이고 사구를 발달시키기 때문에, 갯잔디와 같은 사구식생에게 유리한 환경조건을 조성하였다. 반면에 해홍나물과 퉁퉁마디와 같은 절대염생식물은 지형이 높아짐에 따라 소멸되거나 서식 면적이 급격하게 감소되었다. 또한 갯잔디는 지하경에 의해 식생이 발달하기 때문에 모래와 같은 퇴적물의 이동을 방해하여 다른 염습지 식물보다 군락 내에 퇴적을 가중시키는 기능도 있는 것으로 판단된다. 함평만이 속한 무안갯벌은 세계자연유산 등재를 추진할 만큼 생태적 가치가 높고, 철새도래지로도 중요한 기능을 하고 있다. 이러한 함평만에서 지속적으로 식생과 지형이 변화하고 있는 것이 확인되었다. 함평만 갯벌의 효과적인 자연보전 관리 계획이 수립을 위하여 갯벌의 갯잔디군락의 증가와 해홍나물 등 염생식물 군집의 감소가 향후 갯벌생태계에 미치는 영향에 대해 통합적으로 연구할 필요가 있다.
Acknowledgements
본 연구는 국립생태원 “2022 국가 장기생태연구 (NIE-고유연구(B)-2022-02)” 의 연구비 지원에 의해 수행되었습니다.
Appendix
Appendix 1.
Monthly climate data from 2016 to 2022 of the Mokpo Observatory
|
Year-Month
|
16-01
|
16-02
|
16-03
|
16-04
|
16-05
|
16-06
|
16-07
|
16-08
|
16-09
|
16-10
|
16-11
|
16-12
|
|
Average Temperature (˚C)
|
1.8
|
3.7
|
7.5
|
13.8
|
18.6
|
22.2
|
26.1
|
27.9
|
23.1
|
17.3
|
10.0
|
5.5
|
|
Minimum Temperature (˚C)
|
-1.0
|
0.0
|
3.5
|
9.8
|
14.4
|
19.6
|
23.7
|
24.6
|
20.0
|
14.5
|
6.4
|
1.9
|
|
Maximum Temperature (˚C)
|
5.0
|
7.7
|
12.3
|
18.7
|
23.6
|
25.6
|
29.3
|
32.0
|
27.1
|
20.8
|
14.1
|
9.6
|
|
Average
Precipitation (mm)
|
79.2
|
40.5
|
54.9
|
192.0
|
119.9
|
133.6
|
249.2
|
20.4
|
197.8
|
203.1
|
61.5
|
45.9
|
|
|
Year-Month
|
17-01
|
17-02
|
17-03
|
17-04
|
17-05
|
17-06
|
17-07
|
17-08
|
17-09
|
17-10
|
17-11
|
17-12
|
|
Average Temperature (˚C)
|
2.6
|
3.0
|
6.6
|
13.6
|
18.7
|
21.9
|
27.2
|
27.0
|
22.2
|
16.7
|
9.1
|
2.6
|
|
Minimum Temperature (˚C)
|
-0.6
|
-0.9
|
2.6
|
9.6
|
14.4
|
18.6
|
24.6
|
24.0
|
18.7
|
13.5
|
5.3
|
-0.6
|
|
Maximum Temperature (˚C)
|
6.8
|
7.5
|
11.4
|
18.5
|
23.6
|
26.2
|
30.5
|
30.6
|
26.5
|
20.6
|
13.3
|
6.4
|
|
Average Precipitation (mm)
|
13.9
|
32.3
|
28.5
|
46.5
|
14.0
|
28.5
|
138.2
|
201.8
|
109.4
|
62.7
|
1.4
|
45.3
|
|
|
Year-Month
|
18-01
|
18-02
|
18-03
|
18-04
|
18-05
|
18-06
|
18-07
|
18-08
|
18-09
|
18-10
|
18-11
|
18-12
|
|
Average Temperature (˚C)
|
0.1
|
1.1
|
8.6
|
13.7
|
18.2
|
22.6
|
27.7
|
28.7
|
22.3
|
15.5
|
10.6
|
3.7
|
|
Minimum Temperature (˚C)
|
-2.8
|
-2.1
|
4.4
|
9.6
|
14.7
|
19.4
|
24.8
|
25.8
|
19.3
|
12.0
|
7.0
|
0.8
|
|
Maximum Temperature (˚C)
|
3.3
|
4.8
|
13.5
|
18.3
|
22.4
|
26.9
|
31.5
|
32.8
|
26.1
|
19.6
|
15.0
|
7.1
|
|
Average Precipitation (mm)
|
58.0
|
39.3
|
152.7
|
169.0
|
89.3
|
162.0
|
67.6
|
246.2
|
151.4
|
138.4
|
51.1
|
38.3
|
|
|
Year-Month
|
19-01
|
19-02
|
19-03
|
19-04
|
19-05
|
19-06
|
19-07
|
19-08
|
19-09
|
19-10
|
19-11
|
19-12
|
|
Average Temperature (˚C)
|
2.2
|
3.3
|
7.7
|
12.3
|
18.1
|
21.2
|
24.8
|
27.0
|
22.9
|
17.3
|
10.3
|
4.9
|
|
Minimum Temperature (˚C)
|
-0.7
|
0.2
|
3.8
|
8.5
|
13.7
|
18.2
|
22.5
|
24.3
|
20.1
|
14.0
|
6.6
|
1.9
|
|
Maximum Temperature (˚C)
|
5.5
|
7.1
|
12.4
|
16.7
|
23.1
|
24.9
|
28.0
|
30.5
|
26.5
|
21.3
|
14.4
|
8.1
|
|
Average Precipitation(mm)
|
13.3
|
29.6
|
38.5
|
86.3
|
122.9
|
172.3
|
167.0
|
144.9
|
259.3
|
159.0
|
14.6
|
40.7
|
|
|
Year-Month
|
20-01
|
20-02
|
20-03
|
20-04
|
20-05
|
20-06
|
20-07
|
20-08
|
20-09
|
20-10
|
20-11
|
20-12
|
|
Average Temperature (˚C)
|
3.6
|
4.2
|
7.4
|
10.3
|
17.2
|
22.2
|
23.0
|
27.7
|
21.9
|
15.9
|
11.2
|
3.2
|
|
Minimum Temperature (˚C)
|
1.3
|
0.7
|
3.7
|
6.8
|
13.9
|
19.4
|
21.0
|
25.4
|
19.2
|
12.8
|
8.2
|
0.1
|
|
Maximum Temperature (˚C)
|
6.3
|
7.8
|
11.6
|
14.4
|
21.2
|
25.9
|
25.5
|
30.8
|
25.1
|
19.6
|
14.4
|
6.9
|
|
Average Precipitation (mm)
|
72.0
|
28.9
|
28.8
|
60.9
|
141.4
|
261.5
|
366.3
|
206.4
|
208.6
|
20.5
|
14.0
|
18.4
|
|
|
Year-Month
|
21-01
|
21-02
|
21-03
|
21-04
|
21-05
|
21-06
|
21-07
|
21-08
|
21-09
|
21-10
|
21-11
|
21-12
|
|
Average Temperature (˚C)
|
2.0
|
4.9
|
9.2
|
13.9
|
17.6
|
22.7
|
26.8
|
26.4
|
23.4
|
17.7
|
11.3
|
4.8
|
|
Minimum Temperature (˚C)
|
-1.6
|
1.2
|
5.7
|
10.0
|
13.9
|
19.7
|
24.1
|
23.8
|
20.7
|
14.3
|
7.9
|
1.4
|
|
Maximum Temperature (˚C)
|
5.7
|
8.9
|
13.8
|
18.1
|
21.7
|
26.4
|
30.2
|
29.7
|
26.7
|
21.8
|
15.2
|
8.7
|
|
Average Precipitation (mm)
|
39.7
|
25.9
|
127.4
|
51.9
|
112.3
|
108.3
|
327.5
|
124.4
|
91.4
|
14.7
|
63.9
|
13.6
|
|
|
Year-Month
|
22-01
|
22-02
|
22-03
|
22-04
|
22-05
|
22-06
|
22-07
|
22-08
|
22-09
|
22-10
|
22-11
|
|
Average Temperature (˚C)
|
1.5
|
1.6
|
8.5
|
13.8
|
17.7
|
22.7
|
26.6
|
26.7
|
22.5
|
15.8
|
12.1
|
|
Minimum Temperature (˚C)
|
-1.4
|
-1.7
|
4.6
|
10.0
|
13.8
|
20.0
|
24.0
|
24.6
|
19.5
|
12.4
|
8.3
|
|
Maximum Temperature (˚C)
|
4.8
|
5.4
|
13.2
|
18.9
|
22.6
|
26.1
|
30.0
|
29.4
|
26.2
|
19.8
|
16.9
|
|
Average Precipitation (mm)
|
4.3
|
4.5
|
78.9
|
74.7
|
2.6
|
90.2
|
196.3
|
92.3
|
101.9
|
38.3
|
33.1
|
Appendix 2.
List of halophyte species that appeared in 2016 and 2022
|
Family name
|
Species name
|
2016
|
2022
|
| Chenopodiaceae | Atriplex gmelinii |
●
| |
| Atriplex subcordata |
●
| |
| Chenopodium virgatum |
●
| |
| Kochia scoparia var. littorea |
●
|
●
|
| Salicornia europaea |
●
| |
| Salsola collina |
●
| |
| Salsola komarovii | |
●
|
| Suaeda glauca |
●
|
●
|
| Suaeda maritima |
●
|
●
|
| Plumbaginaceae | Limonium tetragonum |
●
|
●
|
| Convolvulaceae | Calystegia soldanella |
●
|
●
|
| Asteraceae | Artemisia capillaris | |
●
|
| Artemisia fukudo |
●
|
●
|
| Poaceae | Ischaemum anthephoroides |
●
|
●
|
| Leymus mollis |
●
|
●
|
| Phragmites australis |
●
|
●
|
| Zoysia sinica |
●
|
●
|
References
Anderson, S.M., Ury, E.A., Taillie, P.J., Ungberg, E.A., Moorman, C.E., Poulter, B., Ardón, M., Bernhardt, E. S., and Wright, J.P. 2022. Salinity thresholds for understory plants in coastal wetlands. Plant Ecology 223(3): 323-337.
10.1007/s11258-021-01209-2Bang, J.H. 2018. Effects of elevation and herbivores on the distribution of halophytes in a Korean salt marsh Doctoral dissertation, Seoul National University.
Bang, J.H., Joo, S., Lee, E.J., Jeong, S., and Park, S. 2020. Diet of the Mud-Flat Crab
Helice tientsinensis in a Korean Salt Marsh. Wetlands 40: 311-319.
10.1007/s13157-019-01193-4Bernhardt, J.R. and Leslie, H.M. 2013. Resilience to climate change in coastal marine ecosystems. Annual Review of Marine Science 5(1): 371-392.
10.1146/annurev-marine-121211-17241122809195Bray, J.R. and Curtis, J.T. 1957. An ordination of the upland forest communities of southern Wisconsin. Ecological Monographs 27(4): 326-349.
10.2307/1942268Choi, Y.S., Joo, S., Kim, M.S., Han, D., and Jeong, G. 2017. Habitat Use and Food Materials of the Endangered Swan Goose (
Anser cygnoides) during the Wintering Season. Korean Journal of Ecology and Environment 50(2): 266-274. (In Korean)
10.11614/KSL.2017.50.2.266Curtis, J.T. and McIntosh, R.P. 1951. An upland forest continuum in the prairie-forest border region of Wisconsin. Ecology 32(3): 476-496.
10.2307/1931725Etterson, J.R. and Mazer, S.J. 2016. How climate change affects plants’ sex lives. Science 353(6294): 32-33.
10.1126/science.aag162427365440Fan, D., Wang, Y., and Liu, M. 2013. Classifications, sedimentary features and facies associations of tidal flats. Journal of Palaeogeography 2(1): 66-80.
Ganthy, F., Sottolichio, A., and Verney, R. 2013. Seasonal modification of tidal flat sediment dynamics by seagrass meadows of
Zostera noltii (Bassin d'Arcachon, France). Journal of Marine Systems 109: S233-S240.
10.1016/j.jmarsys.2011.11.027Gedan, K.B., Silliman, B.R., and Bertness, M.D. 2009. Centuries of Human-Driven Change in Salt Marsh Ecosystems. The Annual Review of Marine Science 1: 117-141.
10.1146/annurev.marine.010908.16393021141032Jeong, Y.W. 2021. ‘Korea’s tidal flats’ listed as UNESCO World Natural Heritage. KBS news. https://news.kbs.co.kr/news/view.do?ncd=5242400&ref=A. Accessed 13 December 2022. (In Korean)
Kassambara, A. and Mundt, F. 2020. factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R package version 1.0.7, https://CRAN.R-project.org/package=factoextra. Accessed 12 December 2022.
Kil, H.J. and Lee, J.S. 2011. Unrecorded Pulmonate Snail,
Onchidium hongkongensis (Systellommatophora, Onchidiidae) from Korean Waters. The Korean Journal of Malacology 27(3): 191-192.
10.9710/kjm.2011.27.3.191Kim, Y.K. 2022. Established the National Tidal Flat World Natural Heritage Conservation Headquarters in Shinan-gun. KUKMINILBO. https://news.kmib.co.kr/article/view.asp?arcid=0017615111&code=61121111&cp=nv. Accessed 13 December 2022. (In Korean)
Korea National Climate Data Center. 2022. Open MET Data Portal. https://data.kma.go.kr/. Accessed 2 December 2022.
Kroetsch, D. and Wang, C. 2008. Particle size distribution. Soil Sampling and Methods of Analysis 2: 713-725.
10.1201/9781420005271.ch55Kwon, K.K. and Je, J-G. 2002. Preliminary Studies on the Relationship between Reed and Bacterial Communities in the Salt Marsh Environment of Namyang Bay, Korea. Ocean and Polar Research 24(1): 47-53.
10.4217/OPR.2002.24.1.047Lee, C. 2003. Color Korean Plant Encyclopedia (I, II). Hyangmunsa. Seoul, Korea. (In Korean)
Lee, D.I., Tac, D.H., and Kim, G.Y. 2016. Diagnosis of conflict problem between the marine environmental conservation and development, and policy implication for marine spatial planning. Journal of the Korean Society for Marine Environment & Energy 19(3): 227-235. (In Korean)
10.7846/JKOSMEE.2016.19.3.227Lee, H.Y. 2013. Diversity and biomass of benthic diatoms in Hampyeong Bay tidal flats. Korean Journal of Environmental Biology 31(4): 295-301. (In Korean)
10.11626/KJEB.2013.31.4.295Lee, H.Y. and Jung, M.H. 2011. Distribution of benthic diatoms in tidal flats of Hampyeong Bay, Korea. Korean Journal of Environmental Biology 29(1): 17-22. (In Korean)
Lee, K.S. and Jun, S.K. 2009. Characteristics of hydrography and tidal current in Hampyung Bay, the western coast of Korea. Journal of the Korean Earth Science Society 30(2): 247-256. (In Korean)
10.5467/JKESS.2009.30.2.247Lee, S.H., Lee, J.S., Kim, J.W., Kim, H.S., and Ihm, H.B. 1998. Studies on the vegetation distribution and biomass at the wetland of Hampyung-Man. Bull Ins Litt Environ Mokpo Natl Univ, 15: 9-20. (In Korean)
Ministry of Environment. 2020. Korean climate change assessment report 2020 - Adaptation and impacts of climate change. ISBN 978-89-93652-58-1. (In Korean)
National Institute of Biological Resources. 2022. National list of species of Korea. Incheon, Korea. (In Korean)
National Institute of Crop Science. 2014. Food Crop Environment Analysis Method Handbook. National Institute of Crop Science. Incheon, Korea. (In Korean)
National institute of Ecology. 2016. Basic ecological Research for Climate Change Risk Assessment. National institute of Ecology. Seocheon, Korea. pp. 234-256. (In Korean)
National institute of Ecology. 2021. Monitoring ecosystem response to climate change. National institute of Ecology. Seocheon, Korea. pp. 124-146. (In Korean)
Park, H.S., Kwon, B.H., Kim, I.K., So, Y.H., Oh, S.B. and Kang, D.H. 2017. Analysis of Variations in Soil Heat Flux and Air Temperature by Net Radiation at a Mud Flat in Hampyeong Bay. Journal of Environmental Science International 26(9): 1101-1110. (In Korean)
10.5322/JESI.2017.26.9.1101Park, J.S., Lee, H., and Lee. J.S. 2022. Hampyeong Bay Salt Plants and Changes in Soil Environment. Ecology and Resilient Infrastructure abstract book. Seoul, Korea. pp. 9. (In Korean)
QGIS.org. 2022. QGIS Geographic Information System. QGIS Association, http://www.qgis.org. Accessed 20 June 2022.
R Core Team. 2022. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria.
Richards, C.L., Pennings, S.C., Donovan, L.A. 2005. Habitat range and phenotypic variation in salt marsh plants. Plant Ecology 176, 263-273.
10.1007/s11258-004-0841-3Shamsutdinov, N.Z., Shamsutdinova, E.Z., Orlovsky, N.S., and Shamsutdinov, Z.Sh. 2017. Halophytes: Ecological Features, Global Resources, and Outlook for Multipurpose Use. Herald of the Russian Academy of Sciences 87(1), 1-11.
10.1134/S1019331616060083Shin, W., Kim, J.H., and Lee, E.J. 2020. Effect of native
Suaeda japonica structure on the initial seed settlement of an invasive plant
Spartina anglica, Aquatic Botany 161: 103175.
10.1016/j.aquabot.2019.103175So, Y.H., Kang, D.H., Kwon, B.H. and Kim, P.S. 2019. Seasonal variations of CO
2 concentration and flux in vegetation and non-vegetation environments on the Muan tidal flat of Hampyong Bay. Journal of Wetlands Research, 21(4): 257-266. (In Korean)
Yoo, J.W., Kim, C.S., Park, M.R., Jeong, S.Y., Lee, C.L., Kim, S., Ahn, D.S., Lee, C.G., Han, D., Back, Y. and Park, Y.C. 2021. Considerations and Alternative Approaches to the Estimation of Local Abundance of Legally Protected Species, the Fiddler Crab,
Austruca lactea. Journal of Wetlands Research 23, 122-132. (In Korean)
Yook, K. 2015. Application of Linear Programming to the Designation of Marine Protected Areas Considering Biodiversity and Fishery Rights. Doctoral dissertation, Seoul National University. (In Korean)


