

1. 서 론
2. 연구 방법
2.1 조사지 개황
2.2 표본 채집 및 동정
2.3 자료 처리
3. 결과 및 논의
3.1 사주 환경
3.2 곤충상
3.3 군집 구조
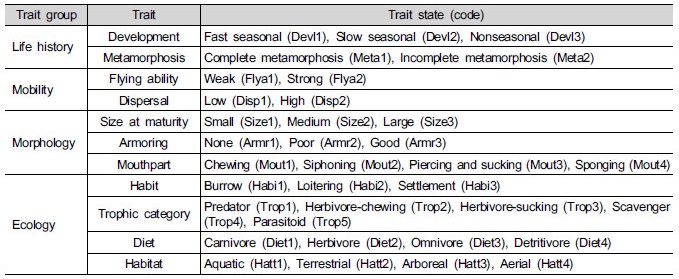
3.4 종 기능군 특성
4. 결 론
1. 서 론
우리나라의 하천은 대부분 인간에 의하여 심각하게 변경, 훼손되어 왔다. 하천정비, 댐 조성, 유역의 토지이용 변화 등에 의하여 물 흐름, 유사의 공급과 이송이 변경되어 하천 생태계 구조와 기능이 교란되었다 (Resh et al. 1988). 이러한 하천 생태계 변화에 사주와 수심이 얕은 서식처의 감소를 포함하는 하도 지형의 변화가 포함된다 (Graf 2006). 더불어 인간 간섭에 따른 물 흐름과 유사 이송의 변화, 수질 오염 등에 의하여 사주 형성이 제한되고 식생이 사주에 번성하게 된다 (Graf 2006, Woo and Park 2016). 사주 형성이 저해되고 기존의 사주에 식생이 고착됨에 따라서 이곳에 정착한 생물상과 군집구조에 변화가 뒤따르게 된다 (Smith 2011, Stucker et al. 2012, Fischer et al. 2015).
유사의 운반이 이루어지는 하천 중류역에서는 일반적으로 모래하천이 형성되고, 이 중류역이 전체 하천에서 가장 길다 (Schumm 1977). 몬순 기후에 영향을 받는 우리나라는 하상계수가 켜서 넓은 사주와 이곳에 하상재료가 노출된 나지 사주가 발달하였다 (Woo 2008). 그러나 하천 공사, 유역 토지이용 변화 등에 의한 유황과 유사 특성의 변화로 나지 사주에 식생이 이입 정착하면서 식생 사주가 확장되고 고정사주로서 육역화되고 있다 (Woo and Park 2016). 모래하천에서 식생사주로 환경이 변화함에 따라서 나지 사주의 고유한 생물상과 생물군집 구조가 점차 변화할 것으로 예상된다.
지구상에는 수백만 종이 넘는 다양한 곤충이 서식하고 있다. 이들의 생태적 특성이 매우 다양하여 먹이망의 다양한 영양단계에서 양적으로 우위를 차지하고 있다 (Gullan and Cranston 2014). 하천에서도 다양한 곤충이 서식하고 있으며 이들이 환경변화에 민감하게 반응하므로 하천 환경에 대한 지표종으로서 이용되고 있다 (Richards et al. 1997, Heino 2010). 하지만 우리나라 하천 사주에서 육상 곤충에 대한 연구는 찾아 볼 수 없다. 단지 하천 사주와 유사한 환경인 해안 사구에서 육상곤충에 관한 연구가 곤충상을 중심으로 수행되었다 (Kim 2003, Han 2010).
낙동강의 1차 지류인 내성천은 대표적인 모래 하상 하천으로서 모래하천 고유의 물리적, 생물적 특성을 유지하고 있다. 내성천의 하상재료는 대부분 중간 내지는 조립질 모래로 구성되어 있고, 식생이 정착한 사주 표면에는 D50이 0.2 - 0.5 mm인 세립질 모래가 퇴적되어 있다 (Lee et al. 2013). 특히 이동상 모래하상이 잘 발달된 내성천에는 연속 만곡부에 소위 백사장인 나지 사주가 형성되어 있다. 이곳의 나지 사주는 회룡포, 무섬마을 등의 역사문화 자원과 연관되어 있다. 그러나 2016년부터 내성천 중류에 건설된 영주댐이 운영됨에 따라서 그 하류 하천에서 급격한 환경 변화와 이에 따른 생물 변화가 예상되고 있다 (Lee et al. 2015).
따라서 본 연구에서는 대표적 모래하천인 내성천의 사주에서 식생 정착에 따른 지면 서식 육상곤충의 생물상과 생물군집의 변화를 파악하기 위하여, 나지 사주와 식생 사주에서 함정법으로 육상곤충을 채집하여 출현 곤충의 종조성, 종다양성, 기능특성군 및 생물군집 구조를 조사하였다. 이러한 연구 결과는 향후 영주댐 운영에 따라서 하류 생태계의 변화를 모니터링하는데 기초선 자료로 이용될 수 있을 것으로 기대된다.
2. 연구 방법
2.1 조사지 개황
본 연구의 대상 하천인 내성천은 경상북도 봉화군 물야면 오전리의 선달산 (1,236 m)에서 발원하여 남류 또는 남서류하면서 영주시, 예천군을 지나 문경시 영순면 달지리에서 낙동강에 유입한다. 내성천의 본류 길이는 111 km, 유역 면적은 1,815 km2이다 (Lee and Kim 2009) (Fig. 1). 내성천은 우리나라의 대표적인 모래 하상 하천으로서 보전가치가 높지만 현재 내성천의 중류에 영주댐이 건설되어 앞으로 급격한 환경 변화가 예상되는 하천이다 (Lee et al. 2015).
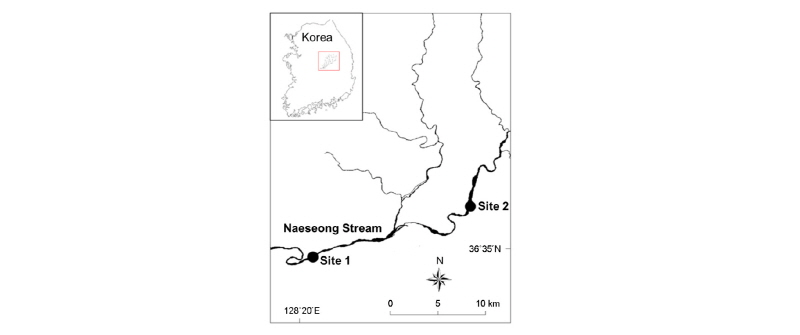
조사 대상 하천인 내성천에서 사주의 식생 정착 정도가 다른 2 구간을 조사지로 선정하였다 (Fig. 1). 각 조사지에서 사주의 제방 방향에서 주수로 쪽으로 트랜젝트를 설치하고 3 조사지점에서 곤충을 채집하였다. 먼저 식생이 유입되지 않은 개방 사주의 조사지는 예천군 지보면 마산리에 위치한 조사지 1 (Site 1)을 선정하였다. 이곳에서는 제방쪽의 고수부지에 달뿌리풀이 정착하여 있지만 물가의 사주에는 식생이 없는 개방 사주가 형성되어 있었다. 이곳에서 제방 쪽의 식생이 있는 곳으로부터 물가로 이동하면서 개방사주에서 조사지점 1-A (Site 1-A), 조사지점 1-B (Site 1-B) 및 조사지점 1-C (Site 1-C)로 나누어 채집지점을 선정하였다. 주로 달뿌리풀이 밀생하여 정착하여 있는 식생 사주의 조사지점은 예천군 호명면 직산리에 위치한 조사지 2 (Site 2)에서 선정하였다. 이곳에서도 제방 쪽에서 물가로 가면서 식생사주에서 조사지점 2-A (Site 2-A), 조사지점 2-B (Site 2-B) 및 조사지점 2-C (Site 2-C)로 나누어 곤충 채집지점을 선정하였다.
2.2 표본 채집 및 동정
조사지의 사주에서 함정 (pit-fall trap) 채집법으로 곤충을 채집하였다. 함정으로는 높이 9 cm, 지름 6 cm의 플라스틱 컵 용기를 이용하였다. 이 함정을 컵의 입구를 지면에 맞추어 사주에 묻었다. 함정 안에는 70% 알코올과 막걸리를 6:4의 비율로 혼합하여 넣었다. 함정은 각 조사지점에서 3개를 5 m 간격으로 설치하였다. 그러므로 식생사주와 나지사주의 조사지에서 각각 12개의 함정을 설치하였다. 함정은 2012년 6월 28일에 설치하고 다음날인 29일에 수거하였다. 수거된 함정 안의 채집물은 플라스틱 용기에 옮겨 실험실로 운반한 후에 분류 동정하였다. 채집된 시료의 동정은 Kim (1970), Lee et al. (1971), Park et al. (2006), Kim (2007), Kwon et al. (2012) 및 Park et al. (2012)를 참고하였다. 동정이 끝난 표본은 액침표본으로 보관하였다.
2.3 자료 처리
내성천의 식생사주와 나지사주에서 곤충의 군집구조를 종다양성으로서 비교하였다. 먼저 각 조사지에서 곤충 군집의 종다양도 지수 (H')와 종균등도 지수 (E')를 산출하였다. 종다양도는 자연로그를 이용한 Shannon의 종다양도 지수를 사용하였다 (Shannon 1948). 종균등도 지수는 군집의 최대 Shannon 종다양도 지수에 대한 실제 종다양도 지수의 비율로서 표시하였다 (Pielou 1975). 또한 군집의 다양성은 순위-수도 곡선 (rank- abundance curve)으로서 표시하였다 (Whittaker 1965). 이 분석은 생물 군집 자료를 이용하여 R 환경 (R Development Core Team 2013)에서 Vegan package (Oksanen et al. 2015)의 ‘radfit’ 함수를 이용하여, Akaike 정보 기준 (Akaike's information criteria, AIC)에 의하여 최적 곡선 모형을 선정하였다.
조사지점 간의 곤충군집 구조를 비교하기 위하여 종별 개체수를 로그로 변환한 입력자료를 이용하여 집괴분석 (cluster analysis)과 서열분석 (ordination)을 실시하였다. 집괴분석은 이용하여 R 환경 (R Development Core Team 2013)에서 Vegan package (Oksanen et al. 2015)의 ‘hclust’ 함수로 계층적 집괴분석을 수행하였다. 거리의 산출은 Bray-Curtis (Bray and Curtis 1957)의 방식을 적용하였다. 집괴분석에서 산출된 높이 수준 (height level)을 기준으로 각 조사지점간 유사거리를 complete 방식으로 분류하였다. 서열분석은 조사지점 사이 거리는 Bray-Curits 방식 (Bray and Curtis 1957)으로 산출하여 비계량형 다차원척도법 (non-metric multi-dimensional scaling, NMDS)를 R 환경 (R Development Core Team 2013)에서 Vegan package (Oksanen et al. 2015)의 ‘metaMDS’ 함수로 수행하였다
사주에서 환경 요인과 곤충의 기능특성 (functional trait)의 관계를 파악하기 위하여 RLQ 분석을 실시하였다 (Dolédec et al. 1996). RLQ 분석을 위하여 곤충 군집의 종별 개체수 (L-matrix), 환경요인 (R-matrix), 출현종의 기능특성군 (Q-matrix)의 3가지 입력 자료를 구성하였다 (Diaz et al. 2008). 환경요인 자료는 조사지점에서 식생 피도, 식생 높이, 수로로부터 거리 및 식생으로 거리로 구성하였다. 출현종의 기능특성은 11개 특성군으로 구분하였다 (Table 1). 구분한 특성군은 생활사로서 발달시기, 변태, 이동성으로서 비행력, 분산, 형태로서 성체 크기, 외장 (armoring), 구기, 생태로서 행동, 영양형, 식성, 서식지이었다. RLQ 분석은 R 환경 (R Development Core Team 2013)에서 ade4 package (Dray and Dufour 2007)의 ‘rlq’ 함수를 이용하였다.
3. 결과 및 논의
3.1 사주 환경
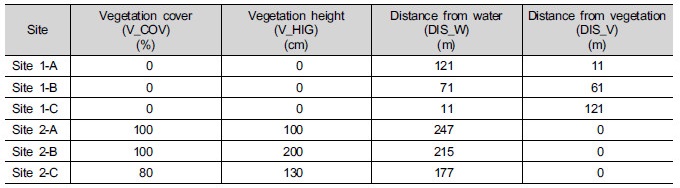
모래하천인 내성천의 사주에서 식생이 정착한 여부에 따라서 선정한 조사지 중에서 조사지 1 (Site 1)은 식생이 정착하지 않은 개방사주이었다 (Table 2). 이 조사지에서 조사지점 1-A (Site 1-A)는 주변 식생으로부터 11 m, 수로로부터 121 m가 떨어진 물가에 가장 먼 곳이었으며, 조사지점 1-C (Site 1-C)은 수로로부터 11 m 거리에 위치한 물에서 가장 가까운 곳이었다. 한편 사주에 식생이 정착한 조사지 2 (Site 2)에서는 3곳의 조사지점에서 식생의 차이를 보였다. 조사지점 2-A (Site 2-A)는 물에서 가장 먼 곳으로서 물쑥 (Artemisia selengensis)과 환삼덩굴 (Humulus japonicus)이 주로 분포하였다. 조사지점 2-B (Site 2-B)와 조사지점 2-C (Site 2-C)에서는 달뿌리풀 (Phragmites japonica)이 우점하였는데, 조사지점 2-B에서는 달뿌리풀이 높이 2 m이었고 매우 밀도가 높았고 조사지점 2-C에서는 달뿌리풀의 밀도가 다소 낮았고 아까시나무 (Robinia pseudoacacia) 등의 다른 식물이 곳곳에 위치하였다.
3.2 곤충상
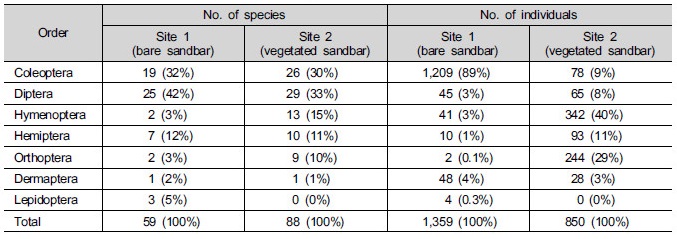
내성천의 나지 및 식생 사주에서 6월에 함정 (pit-fall trap)으로 채집된 곤충은 총 7목 142종 2,209개체이었다 (Table 3). 나지 사주 (Site 1)에 비하여 식생이 정착된 사주 (Site 2)에서 채집된 종수는 많았으나 개체수는 적었다. 종수에서는 식생 사주에서는 특히 딱정벌레목 (Coleoptera), 벌목 (Hymenoptera), 메뚜기목 (Orthoptera)의 종수가 나지 사주보다 많았다. 딱정벌레목은 내륙 산지에서 주로 발견되는 배회성 먼지벌레류가 식생 사주에서 많은 종이 채집되었다. 메뚜기목은 대부분 초식성 혹은 잡식성으로 식생이 침입한 사주에서 더 다양한 종이 분포하였다. 또한 벌목은 주로 맵시벌 종류이었다. 맵시벌은 포식기생자 (parasitoids) 또는 포식자 역할을 수행하며 주로 딱정벌레목과 메뚜기목 등의 유생 또는 성충에 기생하거나 이들을 섭식하므로 딱정벌레목과 메뚜기목이 다양한 식생사주에서 벌목이 더 종수가 많은 것으로 생각된다. 반면에 인시목 (Lepidoptera)은 나지 사주에서만 채집되었으나 4개체에 불과하였다. 내성천 사주의 곤충상을 동해안 사구의 것과 비교하면, 딱정벌레목이 우점하는 것은 유사하지만 사구에 비하여 파리목의 종수가 많았고 노린재목의 종수는 적었다 (Han 2010).
내성천의 나지 사주에서 채집된 총개체수는 식생 사주보다 많았으나, 나지 사주에서는 대부분이 딱정벌레목에 속하였다 (Table 3). 또한 다른 목에서는 식생 사주에서 나지 사주보다 개체수가 많거나 유사하였다. 나지 사주 (Site 1)에서 채집된 딱정벌레목은 대부분이 딱정벌레과 (Carabidae) 먼지벌레류와 풍뎅이붙이과 (His-teridae) 애풍뎅이붙이 종류로서 단단한 외피를 가지면 대부분이 물가에서 채집되었다. 이들 종류는 포식자로서 물가에 모여드는 유충, 사체 또는 쇄설물을 먹이로 섭취하는데 유리할 것으로 생각된다. 또한 나지 사주에서는 모래 환경에 지표종인 것으로 생각되는 강변길앞잡이 (Cicindela laetescripta)와 동양알락방울벌레 (Dianemobius csikii)가 발견되었다 (Lelej et al. 2001, Satoh et al. 2003).
Table 3. The numbers of species and individuals of insects at the bare and vegetated sandbar of the Naeseong Stream. Numerals in the parenthesis indicate the percentages to the total numbers.
 |
3.3 군집 구조
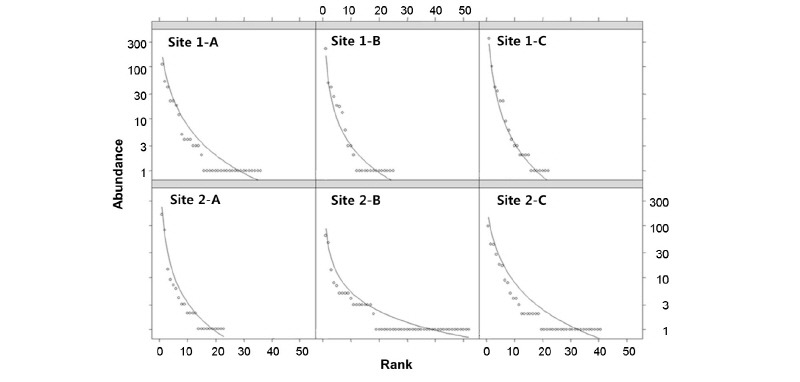
대표적인 모래하천인 내성천 사주에 식생이 침입하면서 곤충 군집의 종다양성 (H')과 종균등도 (E')가 증가하였다 (Table 4). 또한 각 조사지에서 물가로부터 거리에 따라서 곤충의 종다양성을 순위-수도 곡선 (rank- abundance curve)으로 파악하였다 (Fig. 2). 나지 사주에서 저수로로부터 가장 멀고 주변 식생에서 가장 가까운 조사지점 1-A에서 물가의 조사지점 1-C로 갈수록 순위-수도 곡선의 기울기가 급해지고 x 축 절편이 작았다. 따라서 나지 사주에서 주변 식생에서 멀어질수록 종균등도와 종풍부도가 낮아짐을 알 수 있었다 (Whittaker 1965). 반면에 식생 사주에서는 저수로에서 가장 먼 조사지점 2-A에서 종다양성이 가장 낮았다. 이러한 결과를 종합하면 사주에서 식생이 번무하거나 주변 식생과 가깝고 또한 물가에 위치하여 습윤한 곳에서 곤충의 종다양성이 높은 것으로 판단된다.
Table 4. Species diversity (H') and eveness (E') of insect communities at the bare and vegetated sandbar of the Naeseong Stream.
 |
조사지별 곤충 출현종의 수도 자료를 이용하여 수행한 집괴분석에서 나지사주 (Site 1)와 식생사주 (Site 2)의 곤충 군집이 뚜렷하게 구분되었다 (Fig. 3). 식생사주에서는 조사지점 2-B 보다는 조사지점 2-A와 2-B가 서로 유사한 군집구조를 보이고 있었다. 이러한 결과는 비계량형 다차원척도법 (NMDS) 결과에서도 유사하게 나타났다 (Fig. 4). 즉 NMDS 결과에서 1축으로 나지사주와 식생사주가 뚜렷이 구분되었다. 또한 2축으로는 식생사주에서 조사지점 2-B가 2-A 및 2-C와 분리되었다. 조사지점 2-B에서는 큰 키의 달뿌리풀이 밀생하는 곳으로서 달뿌리풀의 피도가 보다 낮고 물쑥, 아까시나무 등의 다른 식물이 침입하여 있는 조사지점 2-A 및 2-C와 다른 곤충 군집의 특성을 보이고 있었다. 이상의 결과를 종합하면 나지 사주에 식생이 침입함에 따라서 곤충 군집의 종다양성이 증가하고 군집 구조도 뚜렷하게 변화하는 것으로 생각된다.
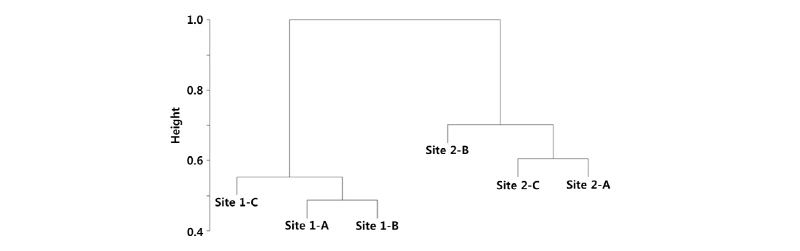
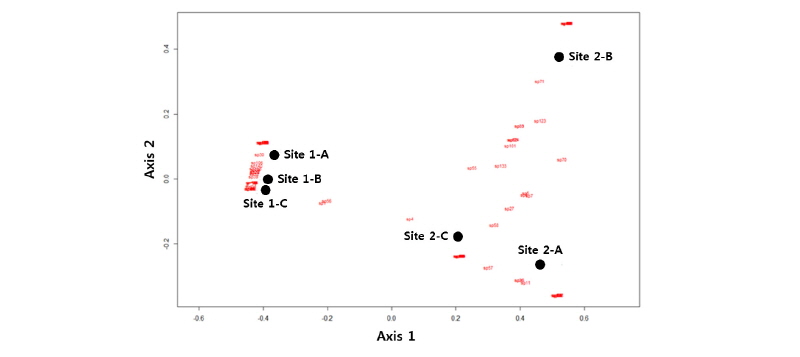
Fig. 4.
Biplot by non-metric multidimensional scaling (NMDS) on the bare (Site 1) and vegetated (Site 2) sandbars at the Naeseong Stream. The alphabets following the site number indicates the positions of the sampling sites: A, far from water channel; B, middle; C, close to water channel. The red characters indicate the insect species.
3.4 종 기능군 특성
내성천 사주에서 곤충 군집 구조에 영향을 미치는 환경요인을 규명하고 곤충 종의 기능특성군의 관계를 파악하기 위하여 RLQ 분석을 실시하였다 (Fig. 5). RLQ 분석에서 1축과 2축의 고유값 (eigen value)이 각각 80.76과 0.06으로서 1축의 설명력이 매우 높았다. 조사지점의 배열에서는 1축의 오른쪽에 나지 사주 (Site 1)가 왼쪽에 식생사주 (Site 2)가 위치하였다. 환경요인과의 관계를 살펴보면 1축의 왼쪽으로 갈수록 식생의 피도와 높이가 높고 물로부터 거리가 멀고 주변 식생과의 거리가 가까웠다. 또한 출현종의 배열과 함께 종의 기능특성군의 배열을 살펴보면 1축에서 식생의 번무하는 왼쪽 방향으로 갈수록 중간 크기 (Size2), 불완전 변태 (Meta2), 초식성(Diet2), 흡즙 초식자 (Trop2), 저작 초식자 (Trop3), 기생포식자 (Trop5)의 특성이 켜졌다. 반면에 나지 사주가 위치한 오른쪽으로 땅굴생활 (Habi1), 완전변태 (Meta1), 포식자 (Trop1), 육식성 (Diet1)의 기능군이 위치하였다. 따라서 사주에 식생이 침입하여 번무함에 따라서 먹이원과 사주 환경이 변화하고 이에 따라서 서식하는 곤충 기능군의 특성에도 변화가 초래되었다.
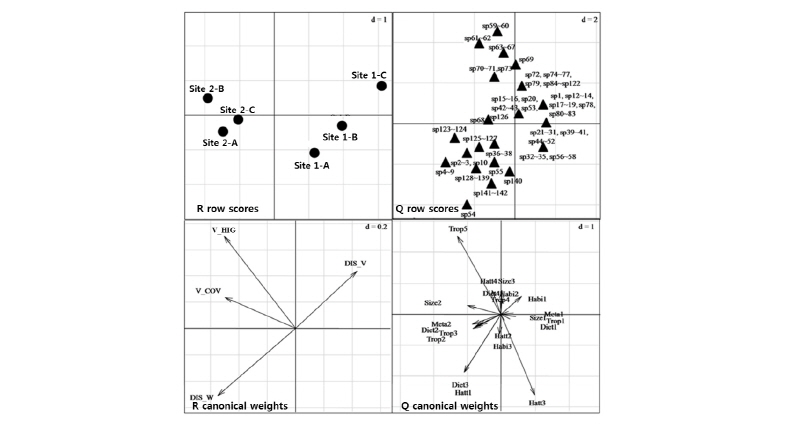
Fig. 5.
Results of RLQ analysis using data on species abundance, environmental variables and functional traits of the insect community at the bare and vegetated sandbars of the Naeseong Stream. Site number refers to Fig. 1 and Table 2, acronym of environmental factors refers to Table 2 and acronym of functional traits refers to Table 1.
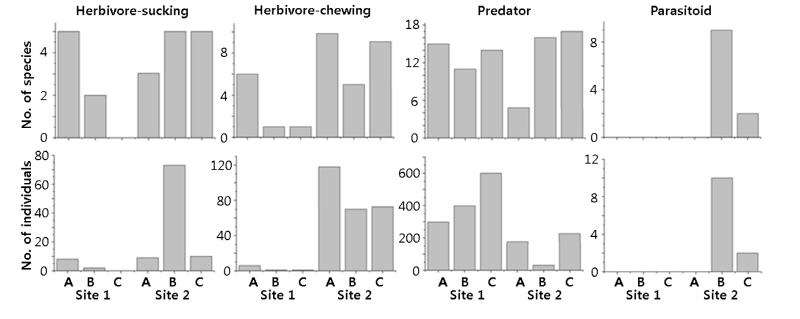
내성천 나지사주와 식생사주의 각 조사지점에서 영양이용형에 따른 기능군별 종수와 개체수를 비교하였다 (Fig. 6). 흡즙 초식자와 저작 초식자의 종수와 개체수는 나주사주 (Site 1)보다 식생사주 (Site 2)에서 많았다. 또한 나지사주에서는 식생에 가까운 조사지점 (Site 1-A)에서 이들이 식생과 먼 조사지점 (Site 1-C)보다 많았다. 포식자는 종수에서는 나지사주와 식생사주에서 비슷하였으나 개체수에서는 나지사주가 식생사주보다 많았다. 반면에 기생포식자는 나지사주에서는 발견되지 않았고 식생사주에서만 관찰되었다. 이들 기생포식자는 식생사주에서도 달뿌리풀이 밀생하는 조사지점 2-B를 선호하였다. 이 조사지점 2-B에서는 흡즙 초식자가 특히 많았다. 이상의 결과를 종합하면 사주에 식생이 번무함에 따라서 초식자 곤충이 증가하고 이에 따라서 포식자보다는 기생포식자의 영양단계가 강화되는 것으로 판단된다.
4. 결 론
모래하천인 내성천에서 사주의 지면에 설치한 함정으로 채집한 지면 서식 곤충의 생물상과 군집구조는 사주에 식생이 침입함에 따라서 먹이원, 서식환경 등의 변화에 따라서 변화하였다. 나지사주에 식생이 침입하여 번성해짐에 따라서 서식 곤충의 종다양성은 증가하였으나 서식 밀도는 감소하였다. 특히 식생사주에서 딱정벌레목, 벌목, 메뚜기목의 종수가 증가하였는데 이들은 하천 사주가 아닌 주변 육상지역에도 서식하는 종들이었다. 나지사주에서는 딱정벌레목의 개체수가 전체 곤충 개체수의 약 90%를 차지하였다. 이들의 대부분은 딱정벌레과 먼지벌레류와 풍덩이붙이과 애풍뎅이붙이 종류로서 건조 환경에 내성이 있고 쇄설물을 먹이원으로 하는 특성이 있었다. 따라서 나주사주에 식생이 침입함에 따라서 종다양성이 증가하지만 나지사주에 특이적이고 희소성이 높은 종의 개체수가 감소하였다.
집괴분석과 서열분석의 결과에 따르면 사주의 곤충군집은 나주사주와 식생사주의 서식지 환경에 따라서 뚜렷하게 구분되었다. 사주의 곤충군집에 영향을 미치는 주요 환경 요인은 사주 식생의 피도와 높이 및 주변 식생과의 거리이었다. 식생 분포에 따라서 곤충 종의 기능군의 특성도 변화하였다. 즉 사주에 식생이 번무함에 따라서 불완전변태, 초식성, 흡즙 및 저작 초식자, 기생포식자 기능군의 비중이 증가하였다. 반면에 나지 사주에서는 완전변태, 포식자, 육식성, 포식자의 기능군의 비중이 증가하였다. 따라서 하천 사주에 서식하는 곤충은 각각의 생태적, 형태적, 기능적 특성에 따라서 자신이 선호하는 서식지에 분포하는 것을 알 수 있었다.
이상의 결과를 종합하여 내성천의 나지 및 식생 사주에서 먹이망을 모식적으로 도시하였다 (Fig. 7). 나지 사주인 조사지 1에서는 에너지 공급원은 주로 유기 쇄설물 (organic detritus)로서 인근 식생이나 홍수터로부터 식물 및 동물 고사체와 분비물 등이 바람과 물에 의하여 공급되는 것으로 생각된다. 이러한 쇄설물은 작은 곤충이나 곤충 유충으로 구성된 쇄설물 섭식자 (detritivore) 또는 청소자 (scavenger)에 의하여 소비된다. 이들은 다시 주로 딱정벌레목 (Coleoptera)에 속하는 포식자에 의하여 피식된다. 반면에 식생이 번무하는 사주에서는 먹이사슬의 바탕이 달뿌리풀 , 물쑥, 환삼덩굴, 아까시나무 등의 1차생산자인 것으로 생각된다. 이들 식물을 초식자인 흡즙 곤충 (sucking insects)와 저작 곤충 (chewing insects)이 이용한다. 조사지에서 흡즙 곤충은 노린재류이었고 저작 곤충은 주로 메뚜기류 이었다. 이들 초식자를 포식하는 상위 소비자는 포식기생자 (parasitoid)와 포식자로서 주로 벌목과 딱정벌레목이었다. 그러므로 모래하천에서 자연 나지 사주에서는 전형적인 쇄설물 기반 먹이사슬을 이루고 있었고, 이곳에 식생이 침입하여 번성하면 점차 초식 먹이사슬로 변하는 것으로 판단된다.
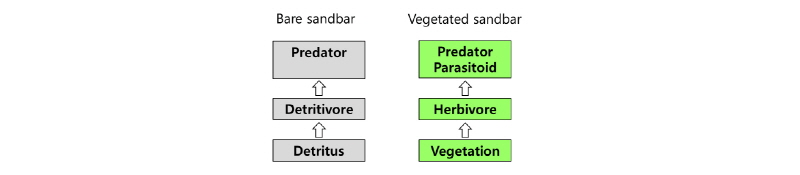
우리나라 모래하천에서 댐 건설, 기후변화 등에 의한 유량과 유사량의 변화, 골재채취, 하천정비, 경작지 홍수터 편입 등과 같은 인위적 교란, 영양물질 유입 증가에 의하여 나지의 사주가 식생이 번무하는 사주로 변화하고 있다 (Woo and Park 2016). 사주에 식생이 정착하고 번무하면 이에 따라서 먹이원과 환경의 변화로 이곳에서 서식하는 곤충 군집의 변화가 본 연구에서 확인되었다. 내성천 나지 사주에는 최근 달뿌리풀, 물쑥, 환삼덩굴, 버드나무, 아까시나무 등의 식생이 점차 번성하고 있다. 이에 따라서 사주에 서식하는 곤충의 종다양성과 서식 밀도가 증가하였다. 또한 종 기능 특성군은 초식 먹이사슬로 변하였다. 식생 사주의 곤충상과 군집구조는 주변 지역의 식생지에서 나타나는 일반적인 유형인 것으로 판단된다. 따라서 나주 사주에서 나타나는 고유성과 희귀성이 높은 쇄설물 기반 먹이사슬을 이루는 곤충 군집과 강변길앞잡이, 동양알락방울벌레와 같은 사주 지표성 곤충상의 보전을 위하여 나주 사주의 보전을 위한 하천 관리가 필요할 것으로 생각된다.
