© Korean Society of Ecology and Infrastructure Engineering.
ABSTRACT
1. 서 론
전 세계적으로 산림생태계의 면적은 인간활동으로 인해 매년 감소하는 추세이다(FAO 2020). 산림생태계는 육상생태계에서 대기 중 CO2를 저감시키는 탄소 저장고로서(Helin et al. 2013, Pan et al. 2024), 2050년까지 탄소중립 목표를 달성하기 위하여 주목해야할 중요한 생태계 유형이다. 전 세계 산림생태계는 지난 20년 동안 약 156억 톤의 CO2를 흡수하는 것으로 계산되었다. 하지만 산림 벌목, 화재 및 인간 활동에 의한 교란 등의 증가로 인하여 연간 81억 톤의 CO2가 숲에서 다시 대기중으로 방출되고 있는 실정이다(Harris 2021). 따라서 더 이상의 탄소 배출을 막고, 산림의 탄소흡수원으로서의 능력을 강화하기 위한 지속적인 노력이 필요하다. 우리나라는 국토 면적의 62.7%를 산림생태계가 차지하고 있다. 전 세계 경향과 유사하게 우리나라의 산림생태계 총 면적은 최근30년(1990년-2020년) 동안 꾸준히 감소하는 추세이다. 하지만 이와 반대로 임목축적량은 꾸준히 증가하고 있다. 2020년 기준으로 우리나라 산림 탄소저장량은 886백만 톤으로 산정되었다(Korea Forest Service 2020).
기후변화에 대비하여 탄소 배출량 감축을 위한 노력은 전 세계적으로 활발하게 진행중이다. 연구자들은 CO2 농도 증가와 온도 상승에 따른 시나리오를 단계별로 가정하였으며(IPCC 2023), 이에 따른 생태계의 반응을 예측하는 시뮬레이션 연구를 활발히 진행하고 있다(National Institute of Environmental Research 2014, Estoque et al. 2019, Liu et al. 2021). 산림생태계 내 수목 동태 변화를 모니터링하고, 기후변화 시나리오에 적용하는 연구는 탄소 동태를 보다 정확히 예측하고 기후 변화와 대응 전략을 수립하기 위한 중요한 기반이 된다.
현장조사 기반의 장기 수집 데이터는 미래 예측 정확도를 높이는데 기여할 수 있다(Gitzen 2012). 특히, 현장에서 수집된 고해상도 수목 생리 데이터는 수목 생장, 군집의 교란 및 기후변화에 따른 반응을 더 잘 이해할 수 있는 좋은 자료이다(Cocozza et al. 2016, Ruehr et al. 2019, Salomón and Camarero 2024). 최근 자동화된 관측 장비를 생태계 모니터링에 적극 활용되고 있으며, 전통적인 방식보다 더 해상도 높은 데이터들이 수집되는 추세이다(Cocozza et al. 2016, Spangenberg et al. 2024). 자동 수목생장측정기(auto dendrometer)는 산림생태계 내 수목의 미세한 생장 변화를 모니터링 할 수 있는 기기로, 간단한 장치를 수목에 설치하여 몇 분 단위의 수목 생장 데이터를 수집할 수 있는 것이 특징이다. 본 연구는 자동 수목생장측정기로 수집된 데이터를 제시하면서, 기존의 전통적인 방법과 비교하고, 이 기기를 활용하기 위한 방법론을 제공하고자 수행되었다. 이러한 연구는 한국 산림생태계의 정확한 탄소수지 산정을 위한 기반이며, 향후 산림의 보전 및 관리 정책 수립에 활용될 수 있다.
2. 연구 방법
2.1 연구 장소
연구장소는 국립생태원 장기생태연구의 일환으로 산림 식생동태 변화 모니터링 연구가 진행되고 있는 강원도 인제군 점봉산, 서울시 용산구 남산, 전남 구례군 지리산의 신갈나무림이다(National Institute of Ecology 2023a). 이 세 지소들은 우리나라 한반도 전체 수준에서 다른 위도에 위치한다. 점봉산 영구방형구의 고도는 800.0±1.2 m, 남산 199.7±0.3 m, 지리산 1365.3±0.3 m이었으며, 각 지소간 고도 차이는 통계적으로 유의했다(Table 1). 각 지소 내 영구방형구는 1-2년 주기로 매목조사가 수행되고있으며, 수고, 흉고직경, 수관활력, 수간건강상태와 같은 정보가 공통적으로 수집되고 있다.
Table 1.
Description of the study sites and the diameters at the breast height (DBH) of Quercus mongolica trees installed with auto dendrometers (mean±SE, n=3). The different letters in the same row indicate the significant difference at p-value <0.05 by Tukey HSD tests
|
Property
|
Site
|
ANOVA
|
|
Jeombongsan
|
Namsan
|
Jirisan
|
F-value
|
p-value
|
|
Location
|
Stand
| Quercus mongolica dominant stand
|
-
|
-
|
|
GPS
|
N38.03813°
E128.46805°
|
N37.55133°
E126.99192°
|
N35.30062°
E127.55282°
|
-
|
-
|
|
Elevation (m)
|
800.0±1.2a |
199.7±0.3b |
1365.3±0.3c |
655322
|
<0.001
|
|
Tree
|
DBH (cm)
|
26.3±1.5a |
26.6±1.3a |
26.2±0.9a |
0.021
|
0.980
|
2.2 대상 수목 선정
연구 대상 수목은 우리나라 산림 대표수종인 신갈나무(Quercus mongoilca)로 선정하였다. 기존 매목조사 자료를 바탕으로 각 지소별로 신갈나무 개체를 3개씩 선별하였다(n=3, N=9). 개체 선별 시 전체 줄기가 곧으며, 병충해 피해나 교란되지 않은 수목인지 확인하고, 수관 활력이 ‘좋음’으로 평가된(1 - 5등급 중 5등급에 해당하는) 개체를 선택하였다. 또한 직경이 23 cm - 29 cm 범위의 수목으로 한정하여 지소 간 비교에서 직경의 영향(즉, 수령의 차이)을 최소화하였다. 결과적으로 세 지소에서 선별된 수목의 DBH는 통계적으로 유의한 차이가 없는 것으로 분석되었다(Table 1).
2.3 자동 수목생장측정기 설치 및 데이터 수집
연구에 사용된 기기는 자동 포인트형 수목생장측정기(Auto point dendrometer, TOMST, Czech Republic)이다(Fig. 1). 이 기기는 수목 직경 변화량을 마이크로미터(μm) 단위로 측정할 정도로 정밀하다. 자동 포인트형 수목생장측정기는 자동 수목생장측정기 종류 중 하나이며, 다른 종류로는 자동 밴드형 수목생장측정기가 있다. 자동 포인트형 수목생장측정기는 밴드형보다 기기 단가가 저렴하고, 설치 및 관리가 간단하므로 여러 수목에 설치할 수 있는 것이 장점이며, 직경 10 cm - 50cm 범위인 수목에 적용하기에 적합하다. 기기 설치 방법은 National Institute of Ecology (2023b)의 기준을 따랐다. 기기는 나무 줄기의 흉고 높이에 나사를 이용하여 고정하고, 압력을 측정하는 센서가 수목 표면에 닿게 세팅 후 하였다. 수목이 수축, 팽창 및 생장함에 따라 센서에 가해지는 압력은 길이로 환산하여 데이터 로거에 저장된다. 초기 설치 시 기기에 가해지는 압력의 환산 값이1000 μm -1500 μm정도가 되도록 하였다. 이는 줄기의 부피가 건조 등에 의하여 수축하였을때도 그 변화를 감지할 수 있도록 하기 위함이다. 또한 기기는 온도센서가 내장되어 있어, 직경 변화량을 기록하는 동시에 연구 현장의 기온 변동도 기록 가능하다. 2023년 여름에 기기를 최초로 설치하였고, 2024년 1월-3월 동안 모든 지소의 세팅을 동일하게 재설정하였다. 데이터 저장 주기는 5분에서 1시간 범위에서 연구자가 원하는 만큼 설정할 수 있다. 10월 말-11월 초 사이에 로거에 기록된 데이터를 다운로드 받았다. 본 연구에서는 2024년 3월 30일부터 2024년 10월 22일까지 207일간(약 7개월) 10분 간격으로 수집된 데이터를 사용하여 연 내 지소별 수목 생장 차이를 추적하였다.

Fig. 1.
View of the automatic point dendrometer installed in the field installation. (a) Photos displaying the installation of a point dendrometer with a contact area adapter and a point pressure sensor. (b) Downloading logged data from the device.
2.4 데이터 분석
10분 간격으로 수집된 데이터에서 정각부터 1시간 간격(정시) 데이터를 추출하고, 일 당 24개 데이터의 평균을 계산하여 일평균 변화량으로 사용하였다. 조사기간 207일 동안의 흉고직경(DBH) 변화로 조사지역별 생장시기와 일일 평균 생장량(cm/day)을 산출하여, Duncan 다중검정을 통해 지소 간 생장량의 차이를 검정하였다. 줄자로 흉고높이의 직경을 측정하는 전통적인 방법을 사용한 결과와 자동 수목생장측정기를 사용한 방법 간 차이를 비교하기 위하여 t-test를 수행하였다. 또한 직경 생장량과 또한 모델 적합성을 기반으로 지역 간 생장 특성을 비교하기 위하여 Day of year를 기준으로 흉고직경 증가 추세를 로지스틱 회귀곡선으로 정량화 하였으며, 이때 사용된 회귀모델식 Eq. 1을 이용하였다(Ajit et al. 1999).
여기에서 a= 최대 생장률(cm), b= 생장 중심 시점, c= 생장속도 이다.
3. 결과 및 논의
점봉산, 남산 및 지리산의 영구방형구에서 신갈나무 수피에 부착된 수목생장측정기로 기록된 2024년 3월 30일 - 10월 22일의 일평균온도는 남산이 20.7±4.9°C(mean±se)로 가장 높았고, 그 다음으로 점봉산 16.2±5.1°C, 지리산 15.0±4.6°C이 가장 낮았다(Fig. 2). 특히 남산은 열섬현상에 의하여 다른 지소에 비하여 기온이 크게 높았고, 위도가 낮은 지리산의 신갈나무림의 기온이 고도가 높아서 위도가 높은 점봉산보다 낮았다. 따라서 수목생장측정기에 설치된 온도 센서로 측정한 온도는 각 조사지의 기온 특성을 잘 반영하고 있어서 온도와 수목 생장 사이의 관계를 파악하는데 유용한 정보를 제공할 것으로 판단되었다.
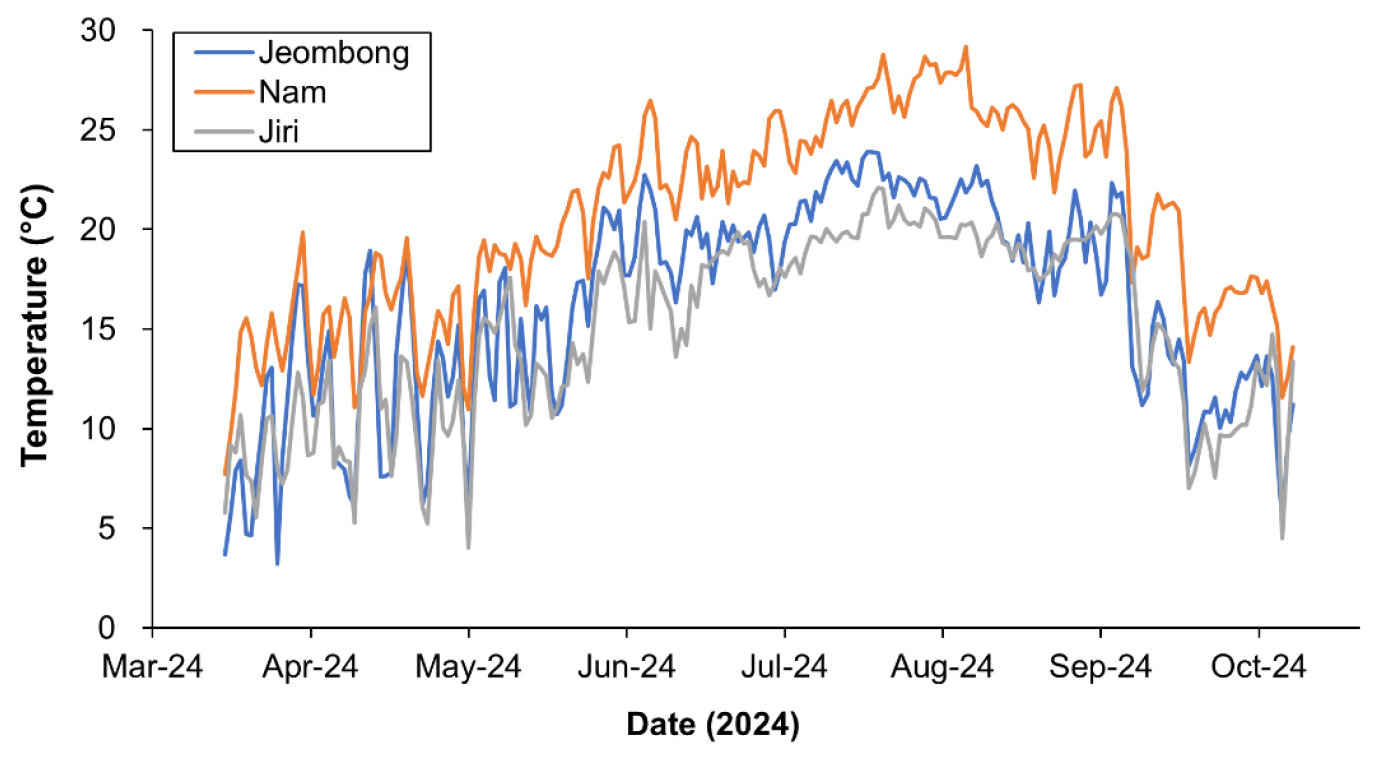
Fig. 2.
Temperature variations in the trunks of Quercus mongolica trees in Mt. Jeombongsan, Mt. Namsan, and Mt. Jirisan, South Korea.
조사기간 동안(2024년 3월 30일부터 10월 22일까지 207일간) 측정한 신갈나무의 일평균직경생장량을 비교한 결과, 지리산이 0.0018 cm/day으로 가장 빨랐고 점봉산과 남산이 각각 0.0013 및 0.0014 cm/day으로 유사한 경향이었으나 이러한 차이가 통계적으로 유의하지는 않았다(Fig. 3 및 Table 2). 그러나, 봄부터 초여름까지의 생육기에 대한 일평균직경생장량을 비교한 결과, 통계적으로 유의한 차이가 있었다. 각 지소의 생육기는 점봉산이 2024년 4월 14일부터 7월 29일까지 107일간, 남산이 4월 5일부터 7월 12일까지 99일간, 지리산이 5월 4일부터 7월 14일까지 72일간이었다(Table 2). 봄철 신갈나무 일평균직경생장량은 지리산이 0.0040 cm/day으로 남산과 지리산보다 통계적으로 유의하게 높았다. 남산과 점봉산은 각각 0.0025 및 0.0022 cm/day으로 유의한 차이가 없었다(Table 2). 지리산 조사지의 신갈나무는 봄철 생장기는 짧지만 이 기간의 일평균 직경생장량이 다른 지소보다 높아서 연간 일생장량도 높은 것으로 판단되었다. 전통적인 방법(수동 측정)과 자동 수목생장측정기로 측정한 방법 간 차이를 비교하기 위하여 t-test를 수행한 결과, 두 그룹간(수동 및 기기) 유의한 차이는 없었다(p-value=0.230).
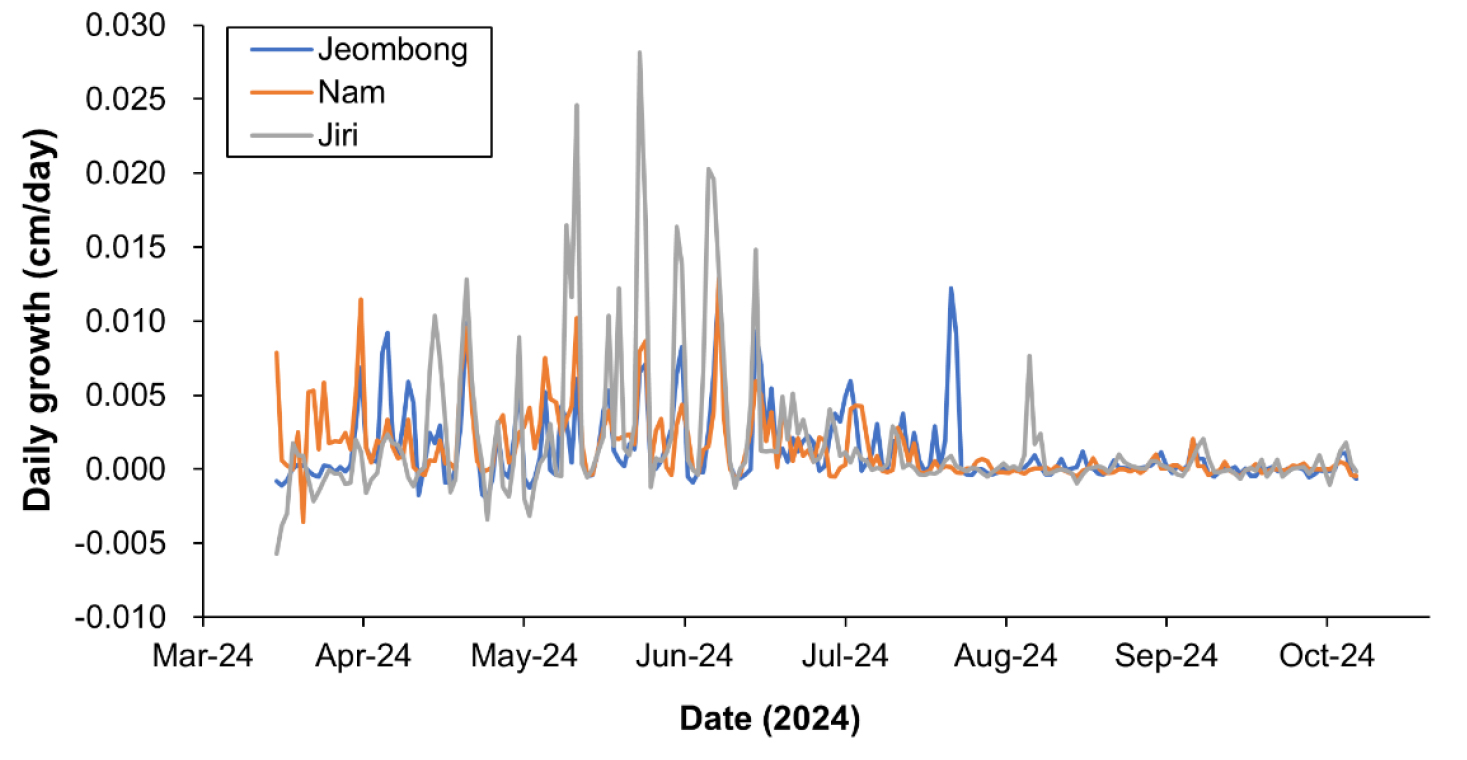
Fig. 3.
Changes of daily growth of the diameters at the breast height of Quercus mongolica in the Mt. Jeombongsan, Mt. Namsan, and Mt. Jirisan, South Korea.
Table 2.
Daily growth (cm/day) the diameters at the breast height (DBH) of Quercus mongolica trees installed with dendrometers in the Mt. Jeombongsan, Mt. Namsan, and Mt. Jirisan in 2024 (mean±SD). The different letters in the same row indicate the significant difference at α = 0.01 by Duncan’s multiple test
|
Period
|
Site
|
|
Jeombongsan
|
Namsan
|
Jirisan
|
Whole season (207 days of
30 March – 22 October)
|
0.0013±0.0025a |
0.0014±0.0023a |
0.0018±0.0046a |
Spring growth season
(Duration)
|
0.0022±0.0029a
(107 days of 14 Apr - 29 Jul)
|
0.0025±0.0027a
(99 days of 5 Apr - 12 Jul)
|
0.0040±0.0068b
(72 days of 4 May - 14 Jul)
|
2024년 신갈나무 영구방형구에서 직경 생장량을 로지스틱 함수로 회귀 분석하여 비교하였다(Fig. 4 및 Table 3). 로지스트 함수는 기온 변화에 따라서 주로 봄철에 이루어지는 수목 직경생장 특성을 잘 반영하였다(Tixier et al. 2020). 신갈나무의 연간 총 직경 생장량(a)의 지소별 지리산이 0.36 cm/yr로서 가장 컸으며 남산과 점봉산이 각각 0.29 및 0.27 cm/yr로서 유사하였다. 또한 신갈나무 직경의 생장 중심 시점(b)은 남산이 DOY 149, 그 다음으로 지리산 DOY 163, 점봉산 DOY 168으로 남산에서 생장이 가장 일찍 시작되는 것으로 파악되었다. 한편 수목 생장 시기의 변화 폭(c)는 점봉산이 24, 남산 21, 지리산 13으로 점봉산 신갈나무의 생장 기간이 가장 길었다. 지리산 신갈나무는 짧은 기간에 집중적으로 생장하는 특성을 보였다. 따라서 자동 수목생장측정기는 기온 특성을 반영한, 수목 생장의 연간동태를 세밀하게 파악할 수 있는 유용한 도구임을 확인할 수 있었다(Sun and Liu 2016).
본 연구에서 지리산의 신갈나무는 평균 온도가 다른 지소에 비해 낮음에도 불구하고 생장량은 가장 크게 증가하는 것으로 조사되었다. 이는 피음이 없는 광조건에서 신갈나무가 다른 참나무 수종들에 비해 빠르게 광합성하는 특징과 연관이 있을 수 있다(Kim et al. 2008). 또한 신갈나무 직경생장에는 생장시기의 기온뿐만 아니라 강수량의 영향도 매우 크다(Lyu et al. 2017). 2024년 1월-10월 월평균 강수량을 비교하면 지리산 135.2±26.8 mm, 남산 124.6±49.9 mm, 점봉산 106.4±31.9 mm이었다. 특히 5월-7월 누적 강수량은 지리산에서 357.7 mm로 점봉산(271.3 mm)이나 남산(274.2 mm)보다 많았고, 이러한 환경요인이 신갈나무 생장에 기여했을 수 있다. 이외에도 충분한 수분과 비옥한 토양 조건(Cheng et al. 2012, Jiang et al. 2015)의 요인도 영향을 미쳤을 수 있다. 어떤 요인이 지소별 신갈나무 생장 차이에 핵심 키(key)인지는 강수량과 상대습도(Meng et al. 2021), 토양온도와 일사량(Biondi et el. 2010), 수액이동(Herrmann 2016)과 같은 추가적인 데이터를 함께 수집하여 분석하고, 식물계절현상 관점이 통합된 연구(Jiang et al. 2015, Kim et al. 2019)를 수행함으로써 더 명확히 할 수 있을 것이다.
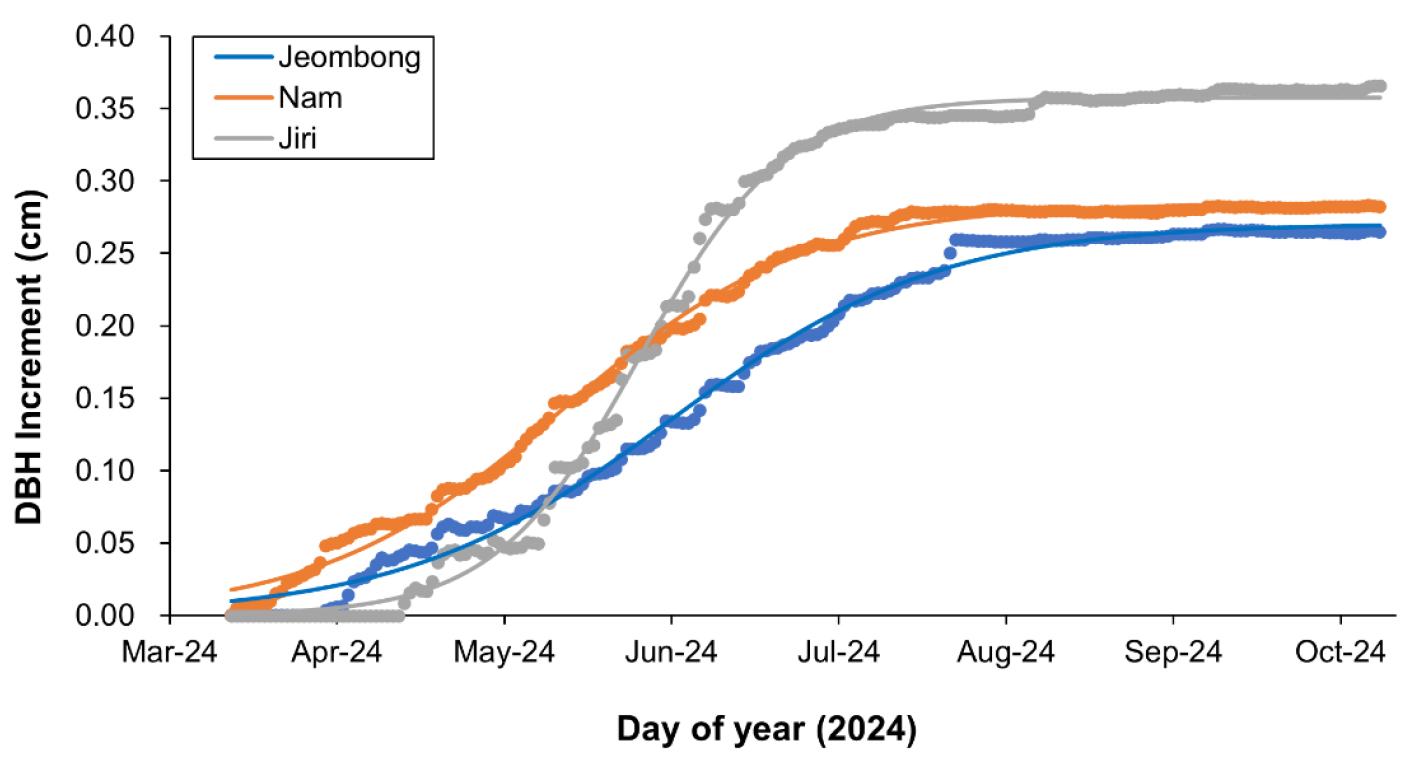
Fig. 4.
Changes of increments of the diameters at the breast height (DBH) of Quercus mongolica in the Mt. Jeombongsan, Mt. Namsan, and Mt. Jirisan, South Korea. The information on the logistic function of the regression line is shown in Table 4.
Table 3.
The coefficient of the logistic regression function† on the increment of the DBH (∆DBH) of Quercus mongolica trees with Day of Year (DOY) in 2024 in the Mt. Jeombongsan, Mt. Namsan, and Mt. Jirisan, South Korea. The regression curve is shown in Fig. 4.
4. 결론 및 제언
자동 수목생장측정기는 고해상도의 시계열 데이터를 축적할 수 있어 수목의 생장 분석에 중요한 도구이다. 이 기기는 정확하고 신뢰할 수 있는 데이터를 제공하기 위해서는 설치 시점과 관리가 매우 중요하다. 가장 적합한 설치 시점은 수목의 생장휴지기인 11월에서 2월 사이이며, 생육기 시작 이후이거나 비가 온 뒤 수피가 수분에 의해 팽창된 시점은 설치를 지양해야 한다. 또한, 수목 줄기가 생장함에 따라 기기의 측정 범위(줄기반경: 10 mm)를 초과할 수 있으므로 1 - 3년마다 정기적으로 기기를 재설치하는 것이 필요하다.
자동 수목생장측정기는 수목 생장 모니터링의 효율성을 높이고, 산림 생태계 관리와 기후 변화 연구에 중요한 역할을 한다. 특히, 고해상도의 연속 데이터를 통해 수목 생장 패턴과 계절적 변화를 정밀히 분석하고, 탄소 흡수 및 배출 동태를 파악하는 데 기여할 수 있다. 그러나 기후변화에 대한 산림의 반응이나 탄소 동태를 이해하기 위해서는 수목의 생리적 변화(광합성, 증발산, 수액흐름 등)와 환경 요인 간 상호작용을 구명하는 것이 우선되어야 한다. 이 연구에 사용된 기기는 온도만을 수집할 수 있으므로, 복잡한 상호작용을 해석하기에는 한계가 있다. 이러한 한계를 극복하기 위해서는 자동 측정 기기를 다양한 환경 조건과 수목 종에 적용하고, 고해상도 미기상 자료, 항공영상, 위성 데이터와 통합 분석을 추진해야 한다. 또한, 기후 변화의 극한 기상 이벤트가 장기적으로 생태계에 미치는 영향을 분석하기 위해서는 다학제적 접근이 요구된다. 이러한 연구는 지속 가능한 산림 관리와 기후 변화 대응 전략 수립에 중요한 통찰력을 제공할 수 있을 것이다.
Acknowledgements
This study was supported by the research grants from the National Institute of Ecology, “2024 Long Term Ecological Research in Korea (NIE-B-2024-02)” and “2024 Integrated Ecological Information Management System for Ecosystem Conservation in Response to the Climate Crisis (2nd) (NIE-C-2024-102)”.
References
Ajit, A., Srivastava, P.N., Gupta, V.K., and Solanki, K.R. 1999. Sigmoid functions in tree growth modelling: limitation of constant estimation and simulation approach as an alternative. Indian Journal of Agroforestry 1(2): 135-140.
Biondi, F. and Hartsough, P. 2010. Using automated point dendrometers to analyze tropical treeline stem growth at Nevado de Colima, Mexico. Sensors 10(6): 5827-5844.
10.3390/s10060582722219689PMC3247734Cheng, X.B., Wu, J., Han, S.J., Zhou, Y.M., Wang, X.X., Wang, C.G., Zhao, J., and Hu, Q.H. 2012. Photosynthesis, leaf morphology and chemistry of Pinus koraiensis and Quercus mongolica in broadleaved Korean pine mixed forest. Photosynthetica 50: 56-66.
10.1007/s11099-012-0005-yCocozza, C., Palombo, C., Tognetti, R., La Porta, N., Anichini, M., Giovannelli, A., and Emiliani, G. 2016. Monitoring intra-annual dynamics of wood formation with microcores and dendrometers in Picea abies at two different altitudes. Tree Physiology 36(7): 832-846.
10.1093/treephys/tpw00926941291Estoque, R.C., Ooba, M., Avitabile, V., Hijioka, Y., DasGupta, R., Togawa, T., and Murayama, Y. 2019. The future of Southeast Asia's forests. Nature Communications 10(1): 1829.
10.1038/s41467-019-09646-431015425PMC6478739Food and Agriculture Organization of the United Nations (FAO). 2020. Global Forest Resources Assessment 2020: Main report (FAO Forestry Paper 197). Key findings, Italy.
Gitzen, R.A. 2012. Design and analysis of long-term ecological monitoring studies. (Ed.), Cambridge University Press, Cambridge, UK.
10.1017/CBO9781139022422Harris, N.L., Gibbs, D.A., Baccini, A., Birdsey, R.A., de Bruin, S., Farina, M., Fatoyinbo, L., Hansen, M.C., Herold, M., Houghton, R.A., Potapov, P.V., Requena Suarez, D., Roman-Cuesta, R.M., Saatchi, S.S., Slay, C.M., Turubanova, S.A., and Tyukavina, A. 2021. Global maps of twenty-first century forest carbon fluxes. Nature Climate Change 11(3): 234-240.
10.1038/s41558-020-00976-6Helin, T., Sokka, L., Soimakallio, S., Pingoud, K., and Pajula, T. 2013. Approaches for inclusion of forest carbon cycle in life cycle assessment-a review. GCB Bioenergy 5(5): 475-486.
10.1111/gcbb.12016Herrmann, V., McMahon, S.M., Detto, M., Lutz, J.A., Davies, S.J., Chang-Yang, C.H., and Anderson-Teixeira, K.J. 2016. Tree circumference dynamics in four forests characterized using automated dendrometer bands. PLoS ONE 11(12): e0169020.
10.1371/journal.pone.016902028030646PMC5193451IPCC. 2023. Summary for Policymakers. In: Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. The Australian National University, Geneva, Switzerland, 1-34.
Jiang, Y., Wang, B.Q., Dong, M.Y., Huang, Y.M., Wang, M.C., and Wang, B. 2015. Response of daily stem radial growth of Platycladus orientalis to environmental factors in a semi-arid area of North China. Trees 29: 87-96.
10.1007/s00468-014-1089-8Kim, E.S., Cho, H.B., Heo, D., Kim, N.S., Kim, Y.S., Lee, K., Lee, S.H., and Ryu, J. 2019. Precision monitoring of radial growth of trees and micro-climate at a Korean Fir (Abies koreana Wilson) forest at 10-minute intervals in 2016 on Mt. Hallasan National Park, Jeju Island, Korea. Journal of Ecology and Environment 43: 1-20.
10.1186/s41610-019-0117-4Kim, S.H., Sung, J.H., Kim, Y.K., and Kim, P.G. 2008. Photosynthetic responses of four oak species to changes in light environment. Korean Journal of Agricultural and Forest Meteorology 10: 141-148.
10.5532/KJAFM.2008.10.4.141Korea Forest Service. 2020. 2020 Forest Basic Statistics. Korea Forest Service. https://kfss.forest.go.kr. Accessed 3 December 2024. (In Korean)
Liu, H., Liu, Y., Wang, C., Zhao, W., and Liu, S. 2021. Landscape pattern change simulations in Tibet based on the combination of the SSP-RCP scenarios. Journal of Environmental Management 292: 112783.
10.1016/j.jenvman.2021.11278334015616Lyu, S., Wang, X., Zhang, Y., and Li, Z. 2017. Different responses of Korean pine (Pinus koraiensis) and Mongolia oak (Quercus mongolica) growth to recent climate warming in northeast China. Dendrochronologia 45: 113-122.
10.1016/j.dendro.2017.08.002Meng, S., Fu, X., Zhao, B., Dai, X., Li, Q., Yang, F., Kou, L., and Wang, H. 2021. Intra-annual radial growth and its climate response for Masson pine and Chinese fir in subtropical China. Trees 35(6): 1817-1830.
10.1007/s00468-021-02152-5National Institute of Ecology. 2023a. 2023 Long Term Ecological Research in Korea. National Institute of Ecology, Republic of Korea. https://www.nie.re.kr. Accessed 3 December 2024. (In Korean)
National Institute of Ecology. 2023b. The Standard Manual for Ecological Monitoring: Tree biomass estimation based on tree inventory survey. Chungcheongnam-do: National Institute of Ecology, Republic of Korea. (In Korean)
National Institute of Environmental Research. 2014. 2013 Annuals of the National Institute of Environmental Research. National Institute of Environmental Research.
Pan, Y., Birdsey, R.A., Phillips, O.L., Houghton, R.A., Fang, J., Kauppi, P.E., Keith, H., Kurz, W.A., Ito, A., Lewis, S.L., Nabuurs, G.J., Shvidenko, A., Hashimoto, S., Lerink, B., Schepaschenko, D., Castanho, A., and Murdiyarso, D. 2024. The enduring world forest carbon sink. Nature 631(8021): 563-569.
10.1038/s41586-024-07602-x39020035Ruehr, N.K., Grote, R., Mayr, S., and Arneth, A. 2019. Beyond the extreme: recovery of carbon and water relations in woody plants following heat and drought stress. Tree Physiology 39(8): 1285-1299.
10.1093/treephys/tpz03230924906PMC6703153Salomón, R.L. and Camarero, J.J. 2024. Stem growth and dehydration responses of Mediterranean tree species to atmospheric and soil drought. Plant, Cell & Environment 26: 832-846.
Spangenberg, G., Zimmermann, R., Küppers, M., and Hein, S. 2024. High-resolution dendrometer measurements reveal different responses of Douglas-fir to extreme drought in 2018 depending on soil and rooting characteristics. Frontiers in Plant Science 15: 1485440.
10.3389/fpls.2024.148544039659421PMC11628273Sun, C. and Liu, Y. 2016. Climate response of tree radial growth at different timescales in the Qinling Mountains. PLoS One 11(8): e0160938.
10.1371/journal.pone.016093827508933PMC4980003Tixier, A., Guzmán-Delgado, P., Sperling, O., Amico Roxas, A., Laca, E., and Zwieniecki, M.A. 2020. Comparison of phenological traits, growth patterns, and seasonal dynamics of non-structural carbohydrate in Mediterranean tree crop species. Scientific Reports 10(1): 347.
10.1038/s41598-019-57016-331941910PMC6962427

