

1. 서 론
2. 재료 및 방법
2.1 식물 재료 및 바이오폴리머 혼합 및 생장 측정
2.2 전해질 유출도 측정
2.3 상대수분함량 (RWC) 측정
2.4 토양 수분함량 분석
2.5 고구마 원형질막 분리
2.6 효소 활성 측정 (enzyme assay)
2.7 Western blot 분석
2.8 통계분석
3. 결 과
3.1 가뭄스트레스 하에서 β-glucan 토양혼합에 따른 고구마의 표현형 비교
3.2 β-glucan 토양혼합 유무에 따른 고구마의 가뭄 스트레스 피해 분석
3.3 β-glucan 토양혼합과 가뭄스트레스에 대한 고구마 잎의 전해질 유출도 및 상대수분함량 분석
3.4 β-glucan 토양혼합에 따른 고구마 PM H+-ATPase 발현 및 활성도 비교
4. 고 찰
5. 결 론
1. 서 론
2023년 현재, 지구의 기후는 기존의 기상예보를 벗어나 예측하기 힘든 극한의 환경을 초래한다. 이에 따라 저온, 고온, 가뭄, 홍수 등의 재난이 발생하며 이러한 환경 스트레스는 농업 생산성에 치명적이다 (Verslues et al. 2023). 환경스트레스에 대한 작물의 생산성 증진 연구는 현재까지도 지속되고 있으며 작물의 유전자 조작을 통한 내생적 (endogenous) 방법과 재배환경을 개선하는 외생적 (exogenous) 방법으로 구분된다 (Arif et al. 2021). 토양에 농업 부산물이나 특정 재료를 혼합하는 외생적 방법은 주로 잡초의 방제나 시비의 용도로 활용되었으며 아직 환경스트레스에 대한 작물생장 증진에는 활용되고 있지 않다. 따라서 다양한 소재를 토양에 혼합하여 작물 생산안정성을 증진시키는 연구가 필요하며 작물의 종에 따른 생리/생태적 반응을 조사하는 것이 중요하다 (Klein and Poverenovc 2020).
최근 바이오폴리머 (biopolymer)를 이용하여 친환경적으로 제방을 강화시키는 연구가 진행되었다. 바이오폴리머를 토양에 혼합하면 토양입자의 응집을 향상시키며 높은 수준의 토양강도를 유지할 수 있었다 (Vahedifard et al. 2015). 친환경 제방에서 자생할 것으로 예측되는 야생종 식물과 일반 작물을 바이오폴리머 혼합 토양에 배양하였을 때 발아 및 생장이 촉진되며, 특히 가뭄 조건에서 생장이 유리한 것으로 밝혀졌다 (Chang and Cho 2012, An et al. 2018, Lim et al. 2018). 한편, 이 연구들은 일반 식물을 대상으로 진행되었으며 괴경이나 괴근을 이용하는 작물에 대한 바이오폴리머의 영향은 연구되지 않았다. 바이오폴리머와 직접 접촉하는 뿌리식물에 대한 연구사례는 심화 연구의 중요한 바탕이며 토양혼합 소재의 선택에 영향을 미칠 것이다.
바이오폴리머는 생물 및 미생물의 생체활동을 통해 생성되는 유기 고분자 화합물을 의미한다. 대표적으로 잔탄검 (xanthan gum), 베타글루칸 (β-glucan), 젤라틴 (gelatin), 키토산 (chitosan) 등이 바이오폴리머로 분류되며 식품 및 의약품 산업 분야에 이용되고 있다 (Chang and Cho 2012). 최근 β-glucan 및 잔탄검을 조합하여 토양처리를 했을 때 토양입자의 응집력 강화 및 식물의 생장을 증진시키는 보고가 있다 (Chang and Cho 2012). 바이오폴리머를 토양에 혼합하면 수분보유력 뿐만 아니라 무기이온의 흡착력이 증진되어 작물을 보호할 수 있다는 보고가 최근 바이오에너지 작물 카멜리나에서 연구되었다. 이 연구들에서 바이오폴리머 토양 혼합에 따른 가뭄 및 고농도의 금속이온 (알루미늄, 아연) 피해 감소 효과가 보고되었다 (Shin et al. 2020, Shin et al. 2021).
식물의 가뭄, 온도, 그리고 중금속 스트레스 환경에서 원형질막 (plasma membrane, PM)은 세포를 감싸고 있는 주요 장벽이며 여러 종류의 원형질막 단백질이 수분 및 이온의 수송을 관할하여 세포의 항상성을 유지시킨다 (Barkla and Pantoja 2010). 이중에서 원형질막 H+-ATPase는 ATP를 소모하여 원형질막 내외의 산도 (pH) 구배를 조성하는 주요 단백질이며, 이를 통해 발생된 구동력 (driving force)는 K+/H+ symporter (HAK, high affinity potassium transporter) 및 Na+/H+ antiporter (NHX, sodium hydrogen exchanger)와 같은 다른 막단백질의 수송활성도에 영향을 미쳐 세포 내부 이온항상성을 조절한다 (Kim et al. 2013, Kim et al. 2019, Zhang et al. 2019). 아쿠아포린 PIP2는 원형질막에서 수분수송을 매개하는 채널 단백질이며, 삼투 변화에 따라 수분을 세포 내외부로 수송하는 통로 역할을 한다. 가뭄 및 고염분과 같은 삼투스트레스 하에서 PIP2는 삼투차이에 의한 수분 유출 통로가 된다. 이를 방어하는 기작으로 PIP2를 원형질막으로부터 내재화시키는 단백질 상호작용과 내포작용이 보고되어 있으며, 이 외에도 인산화 (phosphorylation), 입구개폐 (gating) 등의 기작이 밝혀져 있다 (Kim et al. 2019, Kim et al. 2021, Kim et al. 2022). 따라서 가뭄스트레스 하에서 아쿠아포린의 발현을 확인하는 것은 식물의 피해를 진단하는데 유용하다. 한편, 고구마에서 환경 스트레스에 따른 PM H+-ATPase의 발현 및 활성도 변화에 대한 국내외 연구 사례는 극소수이다. 따라서 국내 고구마 품종에서의 PM H+-ATPase 연구와 외생적 환경 변수를 조절하는 연구는 고구마의 생리 및 생태를 이해하는데 도움이 될 것이다 (Yu et al. 2018, Yu et al. 2019).
본 연구는 바이오폴리머 토양혼합 유무에 따른 고구마의 가뭄스트레스 피해 저감효과를 분석하였으며, 세포 수준에서 환경스트레스 내성에 기여하는 PM H+-ATPase및 PIP2의 발현과 활성을 조사하여 고구마에 미치는 바이오폴리머의 긍정적 효과를 검토하였다.
2. 재료 및 방법
2.1 식물 재료 및 바이오폴리머 혼합 및 생장 측정
본 연구에서 사용된 β-glucan (BIOSYNTH, UK) Table 1에 표기된 비율로 혼합 후 고구마 (소담미) 줄기를 2마디씩 삽식하였다. 삽식한 고구마는 16 h (30°C, 광)/ 8 h (25°C, 암)조건으로 일주일간 생육하였다. 고구마를 생육한 포트에 일정 무게까지 관수 후 급수를 중단하여 가뭄 처리를 하였다. 14일간 급수를 중단한 고구마의 제5엽을 채취하여 엽장 및 엽폭을 측정하였다.
Table 1.
Biopolymer amendment recipe
| Biopolymer concentration (%) | Soil (g) | β-glucan (g) | Water (ml) |
| 0 | 900 | 0 | 270 |
| 1 | 891 | 9 | 270 |
2.2 전해질 유출도 측정
14일간 가뭄 처리한 고구마 줄기에서 제3엽을 각각 0.1 g 수확하였다. 수확한 잎은 4등분하여 30 ml 3차 증류수가 담긴 50 ml conical tube에 넣은 후 26°C, 180 RPM 조건으로 2시간동안 교반 하였다. Blank 값을 측정하기 위해 차 증류수의 전기 전도도 (EC1)를 측정하였고, sample 용액에서 전기 전도도 (EC2)를 IQ170 electrical conductivity meter (IQ scientific instruments, USA)를 이용하여 측정하였다. 측정한 sample들을 모두 고압 멸균 후 상온에서 식힌 후 전기전도도 (EC3)를 측정하였다. 전해질 유출도는 다음과 같은 식을 이용하여 계산하였다.
2.3 상대수분함량 (RWC) 측정
14일간 가뭄 처리된 고구마 제2엽을 채취하여 fresh weight (FW)를 측정하였다. 측정한 잎은 물에 적신 티슈 사이에 포개어 2시간 동안 충분히 침지 하였다. 침지 된 잎에 물기를 티슈로 제거 후 turgid weight (TW) 측정하였다. 측정한 sample은 65°C에서 2일간 완전히 건조시킨 후 dry weight (DW)를 측정하였다. RWC는 아래의 식을 이용하여 계산하였다.
2.4 토양 수분함량 분석
고구마가 삽식된 β-glucan 혼합토양과 일반토양에 충분히 급수하여 수분이 포화된 무게를 측정하였다. 그 후 급수를 중단하여 2 - 3일마다 화분의 무게를 측정하였다. 14일간 무게를 측정한 후 65°C에서 일주일간 완전히 건조시켜 무게를 측정하였다. 토양 수분 함량 측정 식은 아래의 식을 이용하여 계산하였다.
2.5 고구마 원형질막 분리
가뭄 처리된 고구마 지상부를 채취하여 homogenization buffer (pH 7.5, 50 mM MOPs-Bis Tris propane, 330 mM sucrose, 5 mM ethylenediaminetetraacetic acid, 5 mM dithiothreitol, 0.2% bovine serum albumin and 0.5% polyvinylpyrrolidone)를 넣고 얼음위에서 분쇄하였다. 분쇄된 sample은 거즈로 한번 거른 후 4°C, 10,000 g 로 15분간 원심분리 하였다. 상등액을 추출 후 4°C, 80,000 g 로 45분간 원심분리 하여 상등액을 버리고 pellet을 3 ml buffer (pH 7.8, 5 mM KH2PO4, 5 mM KCl 330 mM sucrose, 1 mM ethylenediaminetetraacetic acid and 100 μM dithiothreitol)로 녹여 total membrane을 추출하였다. 추출된 total membrane을 6.5% (w/w) Dextran (MW 45,000 – 65,000), 6.5% polyethylene glycol (PEG 4000), 250 mM sucrose, 4 mM KCl and 5 mM KH2PO4 (pH 7.8)에 잘 섞은 후 4°C, 2000 RPM 10분간 원심분리를 하였다. 그 후 상등액을 채취하여 20 ml final buffer (pH 7.5, 50 mM MOPs- Bis Tris propane, 330 mM sucrose and 5 mM KCl) 섞은 후 4°C, 100,000 g 조건에서 한시간동안 원심분리 하였다. 원심분리된 sample의 pellet을 final buffer에 다시 녹인 후 western blot 및 H+-ATPase 활성 측정 실험에 사용하였다.
2.6 효소 활성 측정 (enzyme assay)
고구마의 원형질막 단백질 샘플들을 각각 30 mM MOPs–BTP (pH 6.5), 3 mM MgSO4, 50 mM KCl, 1.5 mM ATP, 그리고 0.05% Triton-X100으로 조성된 용액에 조심히 혼합하여 30분 동안 37°C에 반응시켰다. 이 반응에서 PM H+-ATPase의 억제제로 200 μM vanadate 그리고 액포의 H+-ATPase 억제제로서 100 mM KNO3가 사용되었다. 그 후 샘플들은 즉시 500 μL 5% trichloroacetic acid 그리고 2 mL 100 mM sodium acetate를 투여하여 반응을 종료시켰다. ATP 가수분해로 인해 발생되는 무기인산을 발색 시키기 위해 300μL 1% ascorbic acid, 60 μL 10 μM CuSO4, 300μ L 0.025 mM H2SO4이 포함된 1% ammonium molybdate 용액을 투여하였다. 10분동안 30°C에서 반응시킨 후 분광광도계 (Biochrome, Libra S50)를 이용하여 720 nm의 흡광도를 측정하였다. PM H+-ATPase의 활성도 계산 방법은 Kim et al. (2013)의 방법과 동일하다.
2.7 Western blot 분석
분리한 원형질막 sample을 Bradford 단백질 정량 분석법 (Bradford 1976)을 이용하여 정량한 10 μg 원형질막 sample을 sodium dodecyl sulfate-polyacrylamide gel을 이용하여 전기영동을 진행하였다. Semi-Dry transfer system을 이용하여 PVDF membrane으로 transfer를 진행하였다. Anti-AHA 항체를 1:1000으로 PBST-20 용액으로 희석하여 membrane을 상온에서 1시간동안 교반 시킨 후 15분간 wash buffer (PBST-20)로 2번 wash 하였다. Horseradich peroxidase가 conjugation 된 Anti-rabbit 항체를 1:4000 PBST-20으로 희석한 용액에서 membrane을 상온에서 1시간동안 교반 후 15분간 wash buffer로 2번 wash 하였다. Membrane을 Claro Sola ECL Solution (Biod, Korea)에 1분간 반응시킨 후 iBright 750 (Invitrogen, USA)를 이용하여 발광을 관찰하였다. 관찰된 단백질의 밀도는 iBright 750 지원 프로그램의 normalization 기능을 이용하여 수치화 하였다.
2.8 통계분석
모든 통계 분석은 SPSS version 20 software (IBM, USA)을 사용하여 수행되었으며 one-way ANOVA 분석을 진행하였으며, 사후검정은 Tukey’s HSD 테스트를 통해 통계분석을 하였다 (P<0.05).
3. 결 과
3.1 가뭄스트레스 하에서 β-glucan 토양혼합에 따른 고구마의 표현형 비교
고구마 (품종 소담미)에서 β-glucan의 토양혼합 효과와 생장 변화를 확인하기 위해 14일 동안 수분공급을 중지하여 가뭄처리를 하였다. 그 결과, 가뭄처리를 하지 않은 대조군에서 β-glucan 혼합에 따른 표현형의 차이를 나타내지 않았다. 가뭄 처리 하에서 β-glucan이 혼합되지 않은 토양은 고구마의 위조 (wilting)가 확인되었지만 β-glucan 혼합 토양의 고구마는 위조를 나타내지 않았다 (Fig. 1 (a)). 이는 고구마 잎의 표현형을 관찰에서도 동일한 결과를 나타냈다 (Fig. 1 (b)). 한편, 엽장과 엽폭의 측정결과 가뭄 처리에 의한 고구마의 생장 저하가 확인되었으나, β-glucan의 혼합 여부는 고구마 잎의 생장에 유의미한 영향을 미치지 않았다 (Fig. 1 (c)). 이 결과는 가뭄에 의한 고구마 잎의 위조가 나타나는 시점이 β-glucan의 토양혼합에 의해 지연되었음을 의미한다.

Fig. 1
Changes in phenotype and leaf growth by β-glucan soil amendment in sweet potato under drought stress. Sweet potatoes were grown in β-glucan amended soil or normal soil. Drought stress was treated by limiting water irrigation for 14 days (a and b). The pictures were taken before and after drought stress. A black bar indicates 1 cm (b). Sweet potato leaf length and width were measured during drought stress (c). Error bars indicating standard errors. The statistics analysis of leaf growth showed no significance between normal and β-glucan soil amendment.
3.2 β-glucan 토양혼합 유무에 따른 고구마의 가뭄 스트레스 피해 분석
일반토양 및 β-glucan이 혼합토양에서 14일 동안 가뭄 처리 후 토양의 수분 함량 변화를 측정하였다. 가뭄 비처리에서 β-glucan의 혼합에 따른 유의미한 변화는 확인할 수 없었으나, 가뭄조건에서 β-glucan 혼합 토양은 처리 2일차 부터 수분 함량이 일반토양에 비해 높게 유지되는 것을 확인할 수 있었다 (Fig. 2). 이는 고구마 재배 시 β-glucan 토양혼합이 토양의 수분 함량을 더 높게 유지시키고 있음을 보여준다.
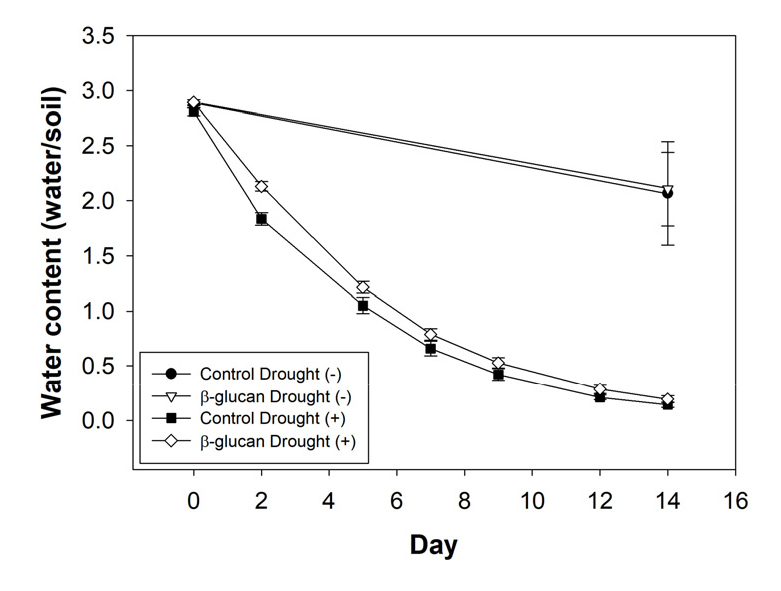
Fig. 2
Changes in soil moisture content by soil amendment with β-glucan under drought stress. Sweet potatoes were grown in pots with soil amended with 1% β-glucan for 14 days with or without water irrigation. Pots with soil were weighed to calculate water content during drought stress. Error bars indicating standard errors.
3.3 β-glucan 토양혼합과 가뭄스트레스에 대한 고구마 잎의 전해질 유출도 및 상대수분함량 분석
가뭄처리 14일 후 고구마 잎을 수확하여 전해질 유출도 및 상대 수분함량을 측정하였다. 가뭄처리 및 β-glucan의 토양 혼합은 고구마 잎의 전해질유출도에 영향을 미치지 않았다 (Fig. 3 (a)). 한편 고구마 잎의 상대 수분함량을 측정하였을 때, 일반토양과 β-glucan 혼합토양에서 생육시킨 고구마 잎은 가뭄 비처리 조건에서 상대수분함량 차이를 나타내지 않았지만, 가뭄 처리 조건에서 β-glucan 혼합토양은 약 20% 높은 수준으로 고구마잎의 수분을 보유하는 것으로 확인되었다 (Fig. 3 (b)). 이 결과는 β-glucan의 토양혼합이 뿌리를 통해 고구마체내로 흡수되는 수분 공급에 유의미한 영향을 주었음을 의미한다.

Fig. 3
Changes in electrolyte leakage and relative water contents of sweet potato leaves by β-glucan amendment in soil under drought stress. The samples were harvested to measure electrolyte leakage (a) and relative water contents (RWC, b) after 14 days of drought stress. Error bars indicating standard errors. Statistical significance was analyzed with SPSS 20 and P<0.01 is indicated by a double asterisk.
3.4 β-glucan 토양혼합에 따른 고구마 PM H+-ATPase 발현 및 활성도 비교
β-glucan의 토양혼합과 가뭄 스트레스에 따른 고구마의 주요 원형질막 단백질 발현 변화를 조사하였다. 그 이유는 가뭄스트레스 하에서 원형질막은 수분의 유출을 방지하고 세포의 항상성을 유지하기 위한 중요한 역할을 하고 있는데, 그 중에서도 PM H+-ATPase 와 아쿠아포린 PIP2는 각각 1차 능동수송 구동력과 수분수송에 중요한 역할을 담당하고 있다. Western-blot 실험을 통해 두 단백질을 발현 변화를 확인한 결과, 가뭄 비처리 조건에서 PM H+-ATPase 및 PIP2의 발현은 모두 β-glucan의 토양혼합에 의해 현저히 증가하였다 (Fig. 4). 또한, 가뭄 처리 조건에서 β-glucan의 토양혼합은 일반토양 샘플에 비해 높은 수준의 PM H+-ATPase의 발현을 나타냈다. 반면, 가뭄 처리 및 β-glucan 토양 혼합에 의한 고구마의 PIP2의 발현이 비혼합토양 샘플에 비해 낮아졌음을 확인하였다. 이 결과는 가뭄 비처리조건에서 β-glucan 토양혼합에 의해 PM H+-ATPase 및 PIP2의 발현이 증가되지만, 가뭄조건에서는 단백질의 종류에 따라 발현의 증감이 다르게 나타날 수 있음을 보여준다.
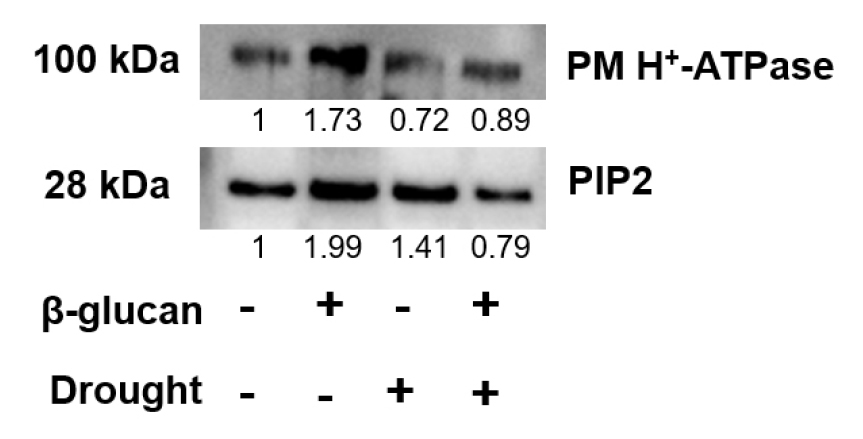
Fig. 4
Western-blot analysis of PM proteins in sweet potato under drought stress. Accumulation of PM H+-ATPase and PIP2 was analyzed by western-blot using against anti PM H+-ATPase and PIP2;1-2 antibodies. The PM samples were collected from shoot of sweet potato grown on normal soil or β-glucan amended soil under drought stress at 14 days. The protein band density was normalized to compare relative accumulation using iBright calculation software.
원형질막 PM H+-ATPase 활성도 변화를 확인하고자 가뭄 처리 후 고구마에서 원형질막 샘플을 추출하였다. 고구마의 PM H+-ATPase characterization test를 실행한 결과, 금속이온 (metal), 계면활성제, ATP의 유무에 의해 PM H+-ATPase 활성도가 각각 55.45 %, 22.69 %, 그리고 93.31 % 감소하였으며, vanadate에 의해 78.18 % 활성도가 감소하였다. KNO3 처리에서 대조구에 비해 92.43 %의 활성도를 나타냈다 (Fig. 5). 이 결과는 고구마의 PM 단백질의 순도 (purity)가 약 80% 이상의 수준으로 활성도 분석의 신뢰도를 확보할 수 있었다.

Fig. 5
Characterization of PM H+-ATPase activity in sweet potato. PM H+-ATPase activity was measured by limited application of metal ion substrates, detergents and ATP or by addition of the ATPase inhibitors vanadate and KNO3. The experiments were triplicated with three individual PM samples. Error bars indicating standard errors.
가뭄 비처리 조건에서 일반토양 및 β-glucan 혼합토양의 고구마 샘플의 PM H+-ATPase 활성도가 유의미한 차이를 나타내지 않았지만, 가뭄 조건 하에서 일반토양 샘플의 PM H+-ATPase 활성도는 비가뭄 샘플에 비해 50% 수준으로 감소하였다 (Fig. 6). 그러나, β-glucan 혼합토양의 샘플은 가뭄처리 하에서 일반토양의 샘플에 비해 약 80% 높은 PM H+-ATPase 활성도를 유지하였다. 이 결과는 β-glucan의 토양혼합이 가뭄 스트레스에 의해 유도되는 PM H+-ATPase 활성의 감소를 지연시키는 효과가 있음을 보여준다.

Fig. 6
PM H+-ATPase activity changes in sweet potato by β-glucan soil amendment under drought stress. The PM samples were collected from sweet potato after 14 days of drought stress. Error bars indicating standard errors. The experiments were triplicated with three individual PM samples. Statistical significance was analyzed with SPSS 20 and P<0.05 is indicated by an asterisk.
4. 고 찰
β-glucan과 같은 바이오폴리머는 농업 생산물 또는 미생물로부터 생산되고 있으며, 바이오폴리머의 생산성 향상 연구들이 수행되고 있다. 하지만 아직까지 농업을 위한 바이오폴리머의 응용 연구는 아직 부족하다. 최근 친환경 제방 건설을 위한 바이오폴리머 응용기술 연구에서 하천 식생에 미치는 영향을 오일작물 카멜리나 및 말냉이와 같은 자생식물에서 바이오폴리머의 영향을 분석한 바 있다 (An et al. 2018, Lim et al. 2018, Jeong et al. 2020). 그 결과, 가뭄 및 중금속 오염 조건에서 바이오폴리머의 수분 보유력 증가 및 금속이온 흡착력이 식물 생장에 유리한 토양 조건을 유지시키는 결과를 확인하였다 (Lim et al. 2018, Shin et al. 2020, Shin et al. 2021). 이 연구들의 연장선에서 바이오폴리머의 종류와 토양혼합 수준, 그리고 스트레스 유형에 따른 고구마의 반응을 분석하고자 본 연구를 수행하게 되었다.
표현형 관찰 실험에서, 일반 토양의 고구마는 가뭄 스트레스에 의한 위조현상이 나타났지만 β-glucan 토양혼합에 의해 위조 현상이 지연됨을 확인할 수 있었다. 하지만 β-glucan의 혼합 유무와 상관없이 14일간의 가뭄 스트레스 동안 고구마의 엽장과 엽폭 생장은 유의미하게 변화하지 않았다 (Fig. 1). 이 결과는 고구마의 생장이 지연될 수준의 가뭄조건에서 β-glucan의 위조점 도달 방지효과를 보여준다. 고구마를 재배하는 토양의 수분 함량은 β-glucan의 혼합에 의해 일반토양에 비해 가뭄조건하에서 높게 유지됨을 보여주었다 (Fig. 2). 이 결과들은 14일 동안의 가뭄처리에서, β-glucan 토양혼합으로 인해 발생된 수분 보유력이 고구마의 가뭄 피해 감소에 도움이 됨을 시사한다.
가뭄 스트레스 피해 수준을 고구마 잎에서 비교하였을 때, 가뭄 처리 및 β-glucan의 혼합 여부는 고구마 잎의 전해질 유출에 큰 영향을 미치지 않았다 (Fig. 3). 그러나 고구마 잎의 상대 수분 함량은 β-glucan의 토양혼합에 의해 유의미한 수준으로 높게 유지됨을 확인하였다 (Fig. 3). 이 결과들은 β-glucan의 토양혼합 기술이 작물의 수분생리에 긍정적인 영향을 미치며 가뭄 스트레스에 대한 방비로서 식생의 보존 및 생산성에 기여할 수 있음을 의미한다.
가뭄 스트레스 하에서 식물의 원형질막은 세포의 수분 및 이온 항상성 유지에 매우 중요하며, 다양한 수송 단백질들이 세포 내외부로 수분, 이온, 그리고 용질 (solute)의 수송을 매개한다 (Kim et al. 2013). 이 중에서, PM H+-ATPase는 1차능동 수송의 구동력을 제공하고, 특히 저온, 가뭄, 이온 스트레스 상황에서 발현과 활성을 조절하여 식물을 생존에 유리한 상태로 전환시킨다 (Kim et al. 2013). 아쿠아포린은 삼투 차이에 따라 세포 내외부로 수분을 수송하는 채널 단백질이며 인산화, 내포작용, 그리고 개폐 조절 기작을 통해 수분수송 활성도를 제어한다 (Maurel et al. 2015). 이러한 원형질막 단백질들의 특성을 근거로 식물의 가뭄스트레스 내성을 평가할 수 있으며, 본 연구에서 수행한 β-glucan의 토양혼합이 고구마 가뭄 스트레스에 대한 피해정도를 평가할 수 있는 생체 지표 (bio-indicator)로 활용하였다.
비가뭄 조건에서, PM H+-ATPase와 PIP2의 발현은 β-glucan 토양혼합에 의해 발현 수준이 높아졌다 (Fig. 4). 이 결과는 β-glucan이 토양의 수분 보유력을 높임과 동시에 주요 원형질막 단백질 발현에 영향을 미치고 있음을 나타낸다. 가뭄 스트레스 조건에서 β-glucan의 토양혼합은 고구마의 PM H+-ATPase의 발현을 가뭄 조건의 일반토양의 샘플보다 더 높게 유지하였고, 이는 PM H+-ATPase 활성도 변화와 유사하게 나타났다 (Fig. 6). 또한, 가뭄처리하에서 혼합토양 고구마 β-glucan 샘플의 PM H+-ATPase 활성도가 유지되는 것은 가뭄 스트레스 피해가 경감되었음을 나타낸다. Fig. 1에서 보여준 영구위조에 가까운 치명적인 가뭄 조건에서 PM H+-ATPase 활성도 변화에 대한 결과는 이 단백질이 세포의 생존력을 나타내는 주요 지표임을 증명해 주었다 (Oh et al. 2014). 저온, 가뭄, 중금속, 그리고, 고염분 스트레스 하에서 식물의 PM H+-ATPase 활성도가 높아지는 것은 잘 알려져 있으며, polyamines와 같은 삼투보호체의 관련성이 보고 되어있다 (Janicka-Russak et al. 2010). 그리고 PM H+-ATPase의 활성에 의한 양자 구동력 (proton motive force)의 영향을 받는 NHX1 단백질의 발현이 고구마의 가뭄 스트레스 내성을 향상시킨 연구가 보고된 바 있다 (Zhang et al. 2019). 이 연구들은 β-glucan의 토양혼합이 고구마의 가뭄 피해 감소에 긍정적 영향을 미침을 명확하게 보여준다. 한편, β-glucan의 영향으로 가뭄스트레스 하에서 상대적으로 발현이 감소한 PIP2는 아마도 내포작용으로 인한 원형질막 내 분포가 감소했을 것으로 추측된다 (Fig. 4). 추가적으로, 카멜리나에서 선행되었던 바이오폴리머 토양혼합에 따른 PIP2;1유전자 발현결과와 고구마의 PIP2 단백질 발현 경향이 동일하게 감소한 것을 확인하였다 (Lim et al. 2018). 이 결과는 극심한 삼투 스트레스하에서 세포 외부로 수분이 유출되는 것을 차단하기위한 PIP2의 내재화 현상과 관련이 있을 것으로 보인다 (Li et al. 2011, Wang et al. 2020, Kim et al. 2021).
