

1. 서 론
2. 연구 방법
2.1 연구 지역과 대홍수
2.2 현장 조사
2.3 이차 효과
2.4 공간 점 패턴 분석 기법
3. 결 과
3.1 회복되지 않은 수목 (“dead”)과 회복한 수목 (“recovered”)의 공간적 관계
3.2 피해를 받은 수목 (“damaged”)과 새로 발생한 수목 (“new”)의 공간적 관계
3.3 회복되지 않은 수목 (“dead”) 및 회복한 수목 (“recovered”)과 새로 발생한 수목 (“new”)의 공간적 관계
4. 토 의
5. 결 론
1. 서 론
전 지구적인 기후변화로 인해 자연적 교란 (natural disturbance)의 빈도, 강도, 지속시간이 증가할 수 있다는 다수의 의견이 존재한다. 홍수 (Meyer et al. 1999), 열파 (Meehl and Tebaldi 2004), 가뭄 (Allen et al. 2015), 산불 (Moritz et al. 2012), 강풍 (Kim et al. 2020a, Webster et al. 2005) 등 세계적으로 교란의 양상 (regime)이 크게 변화하고 있으며, 점차 이에 대비한 새로운 생태계 시스템 관리 방안이 요구된다 (Hannah et al. 2002, Lee et al. In press).
강수가 단시간에 집중할 때 발생하는 홍수는 하천 생태계에 다양한 영향을 준다. 빈번한 주기의 약한 홍수는 식생을 선택적으로 파괴함으로써 군집의 대상분포 (zonation) 양상을 강화시킬 수 있다 (Bendix 1999, Blom et al. 1990). 그리고 이는 종 구성 (species composition) 측면에서의 식생 군집 안정성과도 관련이 깊다 (Bornette 1996). 지형적 측면에서 홍수는 하상물질을 침식시키거나 반대로 신선한 토양을 공급함으로써 미소 서식처 특성을 변화시킬 수 있다 (Stanford et al. 2005, Armstrong et al. 1994). 하지만 기후변화로 인해 발생할 수 있는 대홍수는 그 영향의 양상이 상이하다. 강한 홍수는 하천 지형 구조를 완전히 변화시킬 수 있고 (Eaton and Lapointe 2001), 전역적인 식생 파괴를 초래하여 하천 생태계의 상 전이 (state transition)를 유발할 수 있다 (Poff 2002).
그러므로 대규모 교란이 하천 생태계에 미치는 영향은 대단히 중요한 연구 주제이다. 하천 생태계의 지속 가능한 보전은 생태계 건강성 그 자체와 종 다양성 보전을 위해서도 중요하지만, 홍수시 하천의 수리적 특성이 식생의 영향을 크게 받는다는 점을 고려할 때 (Lee et al. 2021, Makaske et al. 2011) 치수 (治水)의 효율성을 위해서도 중요하다. 국내 하천을 대상으로, 홍수가 하천 생태계에 미치는 영향을 규명하는 연구가 지속적으로 이루어질 필요가 있다 (Kim and Kim 2020, Kim et al. 2020b).
사실 대규모 홍수의 영향을 분석한 연구는 공간적으로 거시적인 스케일에서 진행된 경우가 많았다. 수리모델링을 활용한 홍수 사상 연구 (Roux et al. 2011)나 원격탐사를 활용한 식생 파괴 양상 연구 (Džubáková et al. 2015)를 통해, 대홍수의 영향을 받은 지역을 기술하거나 공간적으로 상이한 피해 양상을 제시할 수 있다. 그러나 미시적 스케일에서 개별 수목을 단위로 진행하는 연구 역시 중요하다. 이러한 접근을 통해 홍수에 반응하는 식생의 변화를 보다 구체적으로 살펴볼 수 있을 것이다.
전 지구적인 기후변화 맥락 속에서 국내에서도 극심한 홍수 발생이 증가할 것이라고 예상되며 (Lee et al. 2004), 이에 따라 변화한 교란 양상에 대응하여 다양한 홍수 취약성 평가 연구들이 진행되고 있다 (see Han et al. 2012, Kang and Lee 2012, Son et al. 2011). 하지만 관련 연구들은 많은 경우 도시 ․ 사회 ․ 경제적 측면에 집중하고 있기 때문에, 대규모 홍수가 국내 하천 생태계에 미치는 영향에 대한 연구가 부족하다고 여겨진다. 이와 더불어 전 세계적으로 자연기반해법 (nature-based solutions)에 중점을 둔 생태계 관리의 필요성이 점차 강조됨에 따라 (Woo and Han 2020) 효과적인 하천 관리를 위해서 하천 생태계 구성 요소들에 대한 구체적인 지식이 요구되고 있다.
본 연구에서는 2020년 여름 한반도 남부를 강타하였던 대홍수로 인해 발생한 수목 피해 양상을 미시적 스케일에서 파악하였다. 더불어 홍수 이후 짧은 시간 동안 비교적 빠르게 발생한 기존 수목 개체의 회복과 새로운 수목의 발생을 공간적 맥락으로 설명하였다. 이를 통해 개별 수목들의 상대적 위치로 대표되는 개체 간 상호작용이 기후변화의 맥락 속에서 가지는 생태적 중요성을 제시하였다. 전통적인 생태학에서 강조해온 환경 변수에 의한 식생 분포의 설명도 중요하지만, 수목 개체 간의 상호작용의 규명 역시 홍수 이후 하천 생태계의 회복을 설명하는데 도움을 줄 수 있음을 강조하였다.
2. 연구 방법
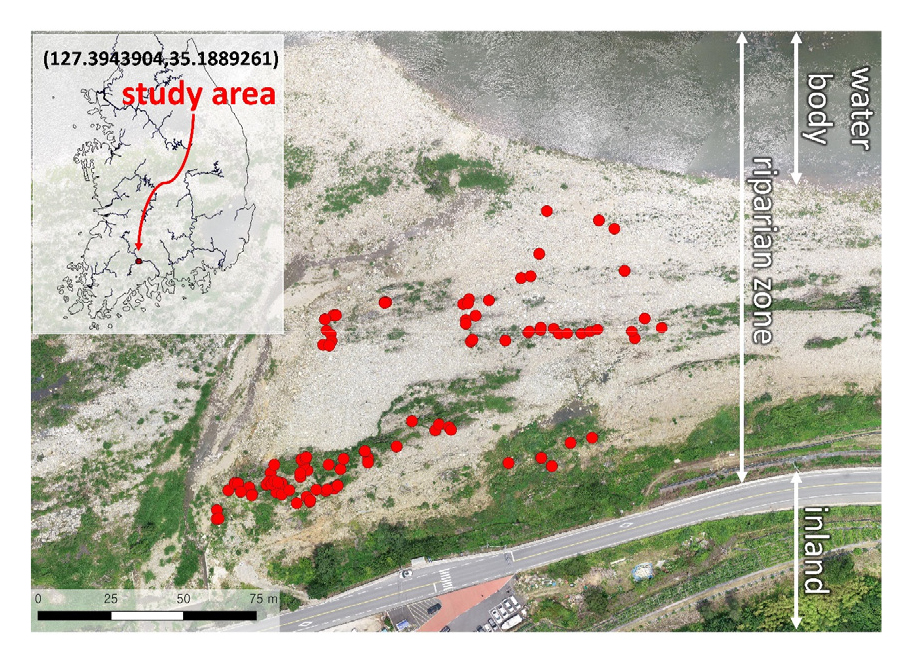
2.1 연구 지역과 대홍수
섬진강은 우리나라 5대강 중 하나로, 한반도 중서부에 위치한다. 팔공산에서 발원하여 전라북도 임실군 신평면에서 그 본류가 시작되며, 섬진강댐을 거쳐 전라남도 광양시 진월면에서 남해로 유출된다. 유로 연장은 222.1 km이고 유역 면적은 4,913.2 km2이다 (MOLIT 2021). 연구 지역은 섬진강 종점으로부터 60.9 km 상류인 전라남도 곡성군 죽곡면 하한리에 위치하고 있는 홍수터이다 (Fig. 1). 연구 지역이 속하는 직선유로의 하상 경사는 1/371 로 중경사를 보이며, 하폭은 271 m이고 만곡되어 있지 않다 (MOLIT 2021). 식생은 주로 버드나무류 (Salix spp.)가 우점해 있었으며, 일부 수변에 갈대류 (Phragmites spp.)가 피복하였고 제방부에 소규모의 왕대림 (Phyllostachys bambusoides)과 경작지가 위치하고 있었다 (Lee et al. 2021).
2020년 여름, 한반도 남부지역을 중심으로 집중호우가 발생하였으며, 본 연구 지역이 위치한 섬진강도 그 영향권에 포함되었다. 특히 섬진강 유역 내 순창 관측소에서 2020년 8월 7 - 8일 양일 간 515 mm의 강수가 관측되었고, 이는 1963년 이래 최대 기록이었다 (HRFCO 2021). 연구 지역의 1.8 km 상류에 위치한 예성교 관측소에서 관측된 대홍수 시 최대 수위는 EL. 43.1 m로, 2020년 평수위인 EL. 28.5 m보다 약 15 m 높았을 뿐만 아니라 해당 지역 계획 홍수위인 EL. 39.5 m도 크게 상회하는 수치이다 (HRFCO 2021). 이로 인해 본 연구 지역의 식생 피복 면적 중 약 80%가 훼손되었다고 보고된 바 있다 (Lee et al. 2021). 한편 해당 대홍수 이후부터 본 연구를 위한 현장 조사가 이루어진 날까지 최대 수위는 EL. 29.3 m로 (MOE 2022), 연구지역에서 추가적인 홍수 교란이 발생하지 않았다고 보아도 무방하다.
2.2 현장 조사
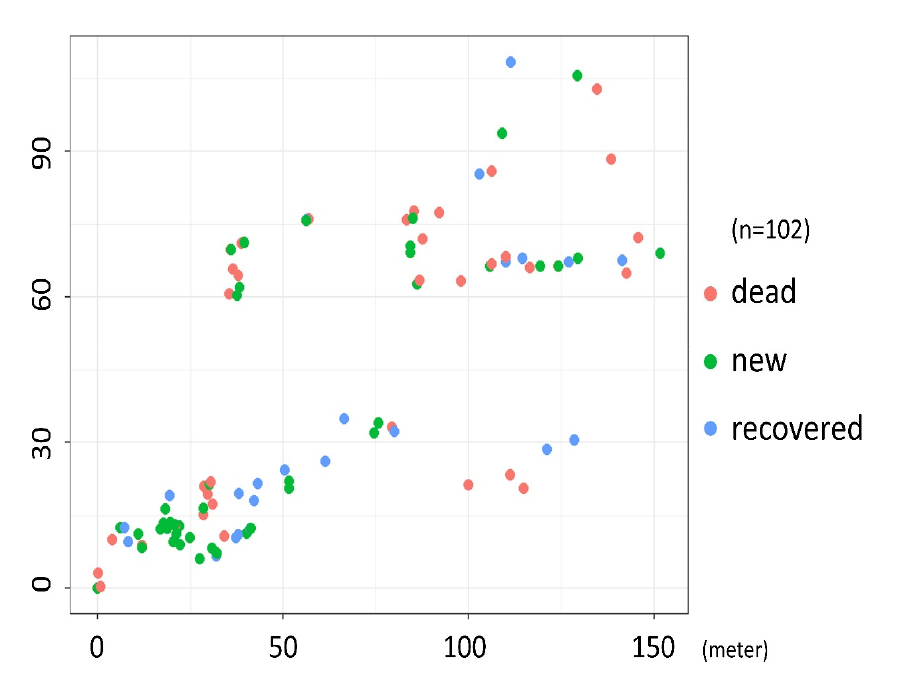
대홍수 발생 후 약 10개월이 지난 2021년 6월 4일, 연구 지역 내 약 1 ha 구역에 분포하는 수목을 조사하였다 (Fig. 1). 개별 수목의 정밀 3차원 좌표를 Real-Time Kinematic (RTK) GPS (model: Trimble R2, 수직 오차: 20 mm, 수평 오차: 10 mm)로 취득하였고, 각 개체의 특성을 세 가지로 분류하였다 (Fig. 2). 첫 번째는 대홍수로 인해 피해를 입은 후 “회복되지 않은 (dead) 수목”으로서, 뿌리는 지면 아래에 여전히 박혀 있지만 조사 시점이 초여름이었음에도 불구하고 잎이 하나도 없는 수목이 해당되었다. 두 번째는 대홍수로 인해 피해를 입었으나 어느 정도 “회복한 (recovered) 수목”으로서, 홍수로 인해 피해를 입은 것은 명백하나, 기둥에서 새로운 줄기가 뻗어 나와 새 잎이 돋아난 수목이 해당되었다. 마지막은 “새로 발생한 (new) 수목”으로서, 홍수의 피해를 받은 흔적이 없고 수직 높이가 1 m 이상인 어린 수목이 해당되었다. 추가적인 분석을 위해, 앞서 언급한 “회복되지 않은 수목”과 “회복한 수목”을 합쳐 대홍수의 “피해를 받은 (damaged) 수목”으로 합쳐서 분류하기도 하였다.
본 현장 조사에서는 개별 수목에 대해 종 단위까지 동정이 이루어지지 않았기 때문에 자료의 세밀함 측면에서 어느 정도의 한계가 존재한다. 그러나 동일한 연구 지역에서 진행된 선행연구 (Lee et al. 2021)에 따르면, 홍수 직후 관찰된 피해 수목은 대부분 버드나무류 (Salix spp.)였다. 아울러 본 연구의 조사 중에 파악한 “회복한 수목”도 대부분 버드나무류였기 때문에, 연구 결과를 해석함에 있어서 큰 어려움은 없을 것으로 판단되었다.
추가적으로, 홍수 이전 개별 수목의 공간적 분포 자료가 존재하지 않아 대홍수로 인해 뿌리째 뽑혀 떠내려간 개체를 고려하지 못한다는 한계가 있다. 홍수시 물에 떠서 빠르게 이동하는 유목 (流木)은 유수 (流水)와 더불어 수목을 파괴하는 또 다른 요인이며 (Sigafoos 1964), 이동하다 하도에 퇴적되어 지형을 변경하기도 한다 (Kang et al. 2020). 뿌리째 뽑힌 수목 역시 대홍수의 피해를 입은 개체라는 점에서 자료를 구득할 수만 있다면 향후 연구에서 고려해야 할 요인이다.
2.3 이차 효과
교란으로 인해 변화하는 식생 군집을 연구할 때, 기본적으로 환경적 맥락을 설명 변수로 투입할 수 있다. 이러한 연구 방법론은 수리적 특성, 지형 조건 등 거시적인 환경 조건이 공간적 변이를 보임에 따라 달라지는 식생 반응을 보여줄 수 있으며, 이를 통해 일차 효과 (first-order effect)를 규명할 수 있다. 상관관계 분석, 회귀 분석, 종 분포 모형 (species distribution model) 구축 등이 그 예시이다. 하지만 환경 변수뿐만 아니라 식물 개체 간의 생물학적 ․ 비생물학적 상호작용 역시 교란으로 인한 식생 군집의 변화를 설명하는 데 유용하다. 공간적으로 인접한 개체일수록 더 긴밀한 상호작용을 맺는다는 가정을 기반으로, 개별 수목의 상대적 위치를 통해 식생 분포를 설명할 수 있다 (Ben-Said 2021, Stoyan and Penttinen 2020). 이는 교란에 대한 식생의 반응을 이차 효과 (second-order effect)로 설명하는 방법이며, 대표적인 기법으로는 후술할 공간 점 패턴 분석 (spatial point pattern analysis)이 있다.
2.4 공간 점 패턴 분석 기법
공간 점 패턴 분석은 공간 상에 점으로 나타낼 수 있는 객체들 (features)의 상대적 위치에 대한 공간 분석 기법이다. 생물지리학과 생태학 등 다양한 분야에서 특히 수목 분포 자료를 통해 생태적 과정을 규명하는 연구에 널리 사용되고 있다 (Ben-Said 2021, Kim et al. 2020a, Stoyan and Penttinen 2000). 이 기법을 통해 동일한 특성을 가진 점 객체들의 분포 양상 (unmarked point pattern)을 군집 (clustered) 분포, 무작위 (random) 분포, 그리고 규칙적 (regular) 분포로 구분할 수 있다. 더 나아가 서로 다른 분류군에 속하거나 연속 변수를 속성값으로 가지는 점들의 분포 양상 (marked point pattern)을 서로 군집하는 (attracted) 분포, 서로 독립적인 (independent) 분포, 그리고 서로 밀어내는 (repulsed) 분포로 구분할 수도 있다. 여기서 특징적인 것은 상이한 공간적 스케일에서 서로 다른 분포 양상을 파악할 수 있다는 점이다. 예를 들어, 연구 지역 전체의 스케일 (넓은 공간적 스케일)에서 개별 수목이 서로 무작위적 분포를 보이더라도, 패치 (patch) 스케일 (좁은 공간적 스케일)에서는 군집 분포를 보일 수도 있다. 공간 점 패턴 분석은 이러한 다중스케일적 접근을 가능케한다.
본 연구에서는 개별 수목의 공간적 분포를 정량화하기 위해 공간 점 패턴 분석 기법의 일종인 K-함수 (Ripley’s K-function)와 g-함수 (pair correlation function) 계열 기법들을 사용하였다. 전자가 특정 점으로부터 일정 범위 내에 있는 점들을 모두 고려하여 통계량을 산출한다면, 후자는 특정 점으로부터 일정 거리만큼 떨어진 점들 만을 고려하여 통계량을 산출한다 (Fig. 3). 즉 K-함수는 어느 정도의 공간적 스케일부터 점 분포가 통계적으로 유의미한 패턴을 이루는지 제시할 수 있으며, g-함수는 통계적으로 유의미한 점 분포 패턴이 탐지되는 특정한 공간 스케일을 제시할 수 있다. 이와 관련하여, g-함수와는 달리 K-함수 분석에서는 좁은 공간적 스케일에서의 분포 양상이 넓은 공간적 스케일에서의 분포 양상에 영향을 미칠 수 있기 때문에 해석 상의 주의가 필요하다.
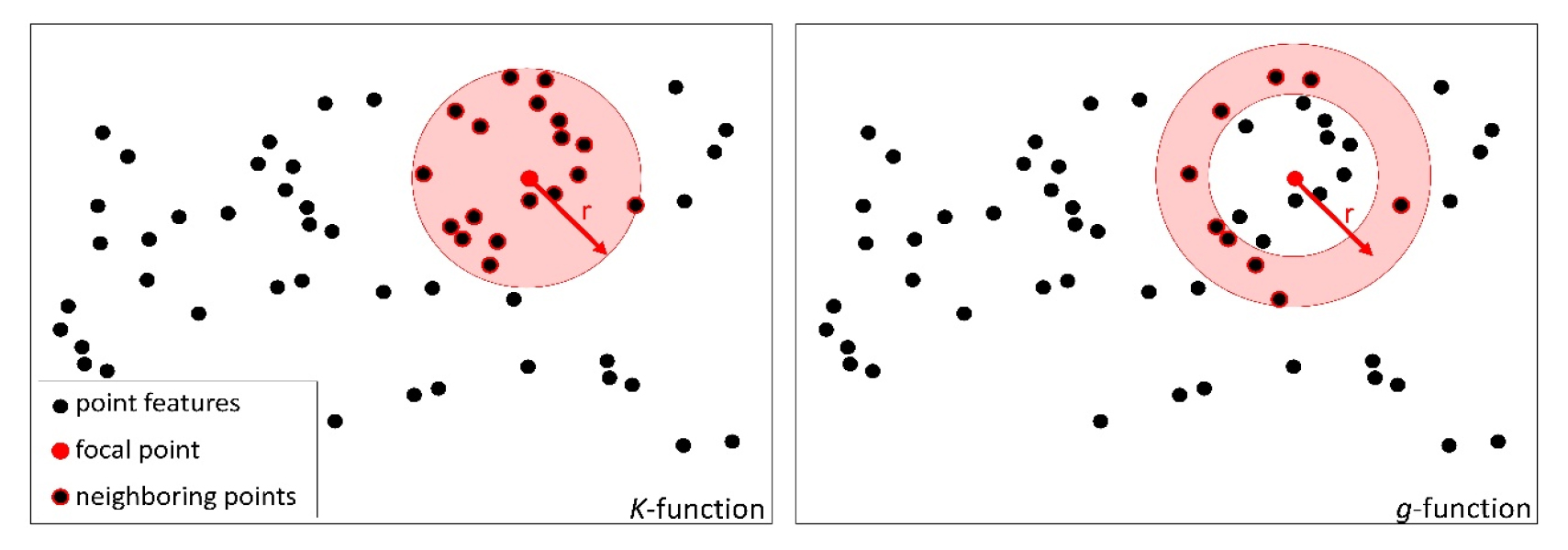
Fig. 3
The difference between the K- and g-functions. The K-function searches the number of neighboring points inside a circle. The count increases cumulatively as the spatial scale (r) increases. In contrast, the g-function searches the number of neighboring points falling within a narrow distance band.
현장 조사 시 연구 대상 수목의 특성을 세 종류 (“dead”, “recovered”, “new”)로 구분하였기 때문에, 조사의 결과는 서로 다른 분류군에 속하는 점 자료라고 볼 수 있다. 이에 따라 서로 다른 두 종류의 수목에 대한 분석을 위해 세부적으로는 K-함수 계열의 bivariate marked Ripley’s K-function과 g-함수 계열의 bivariate marked pair correlation function을 사용하였다 (see Ben-Said 2021). 이를 통해, 하천 수목 전체의 공간적 분포 양상보다는 서로 다른 특성의 개별 수목이 상대적으로 어떻게 분포하는지 정량적으로 밝히고자 하였다. 9,999번의 무작위 순열 검정 (random permutation test)을 통해 통계적 유의성을 시각적으로 제시하였다. 모든 분석은 R 4.0.5 버전에서 진행되었으며, “spatstat” 패키지 (Baddeley et al. 2015)의 “Kcross”와 “pcfcross” 함수가 활용되었다.
3. 결 과
조사 결과, 총 102개 수목 중 “회복되지 않은 수목 (dead)”은 36개 (35.3%), “회복한 수목 (recovered)”은 24개 (23.5%), “새로 발생한 수목 (new)”은 42개 (41.2%)였다 (Fig. 2). 그러므로 대홍수의 “피해를 받은 수목 (damaged)”은 조사된 수목의 약 58.8%를 차지하는 것으로 파악되었다.
3.1 회복되지 않은 수목 (“dead”)과 회복한 수목 (“recovered”)의 공간적 관계
우선, 대홍수의 “피해를 받은 수목 (damaged)” 중 회복 여부가 이차 효과로 설명되는지 규명하기 위해 “회복되지 않은 수목 (dead)”과 “회복한 수목 (recovered)”에 대해 공간 점 분포 분석을 실시하였다 (Fig. 4). 분석 결과, 대부분의 공간적 스케일에서 두 종류의 수목의 상대적 위치는 서로 독립적인 것으로 파악되었다. 비록 K-함수 분석 결과 12 m 이상의 스케일에서 약하게 서로 군집함이 밝혀졌지만, 이는 g-함수 분석 결과에서 나타난 12 m 스케일에서만의 군집성의 영향인 것으로 판단된다. 본 연구 결과에서 주목할 만한 점은 바로 좁은 공간적 스케일에서는 통계적으로 유의미한 군집성이 관찰되지 않았다는 것이다. 즉 수목 패치 (현장 관찰 결과, 대략적으로 r < 10 m) 내 상대적 위치로는 홍수 이후의 회복 여부를 설명할 수 없었다.
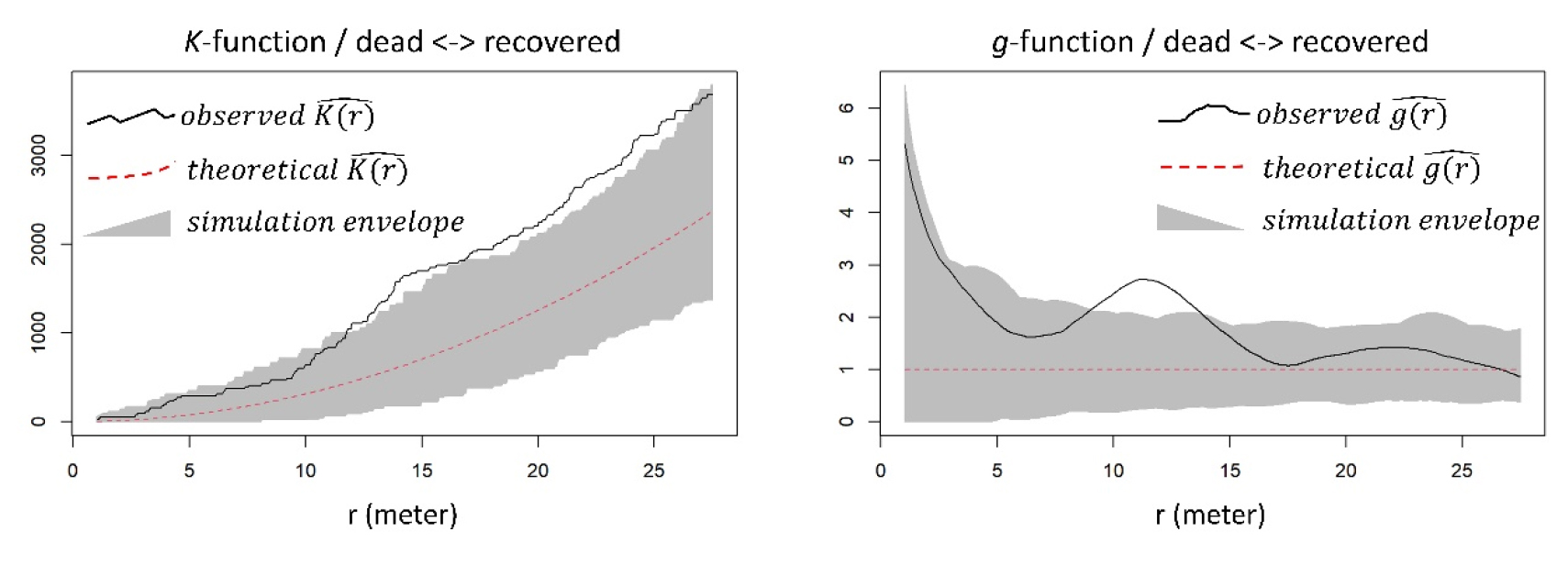
Fig. 4
Bivariate marked spatial point pattern analysis for dead and recovered trees. The observed values located above the envelope indicate a statistically significant attracted pattern. The observed values located inside the envelope indicate an independent pattern. The observed values located below the envelope indicate a statistically significant repulsed (regular) pattern. The horizontal axis shows different spatial scales of the pattern detection. (left) bivariate marked Ripley’s K-function (right) bivariate marked pair correlation function.
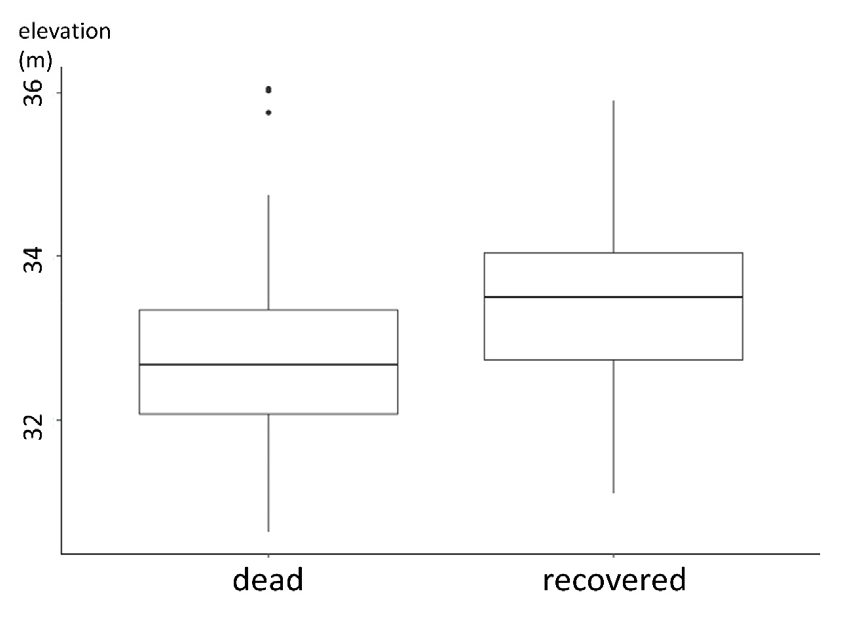
다만, 제한된 자료를 통한 일차 효과 분석에서 회복 여부를 일부 설명할 수 있었다. 수목 위치의 좌표를 취득하는 과정에서 함께 수집된 정밀 고도 자료를 바탕으로 “회복되지 않은 수목”과 “회복한 수목”이 위치한 고도의 평균을 비교하였다 (Fig. 5). 비모수 평균 비교 기법인 Mann-Whitney U 검정 결과 “회복한 수목”이 “회복되지 않은 수목”보다 고도가 더 높은 곳에 위치하였으며, 이는 p < 0.10 수준에서 유의했다.
3.2 피해를 받은 수목 (“damaged”)과 새로 발생한 수목 (“new”)의 공간적 관계
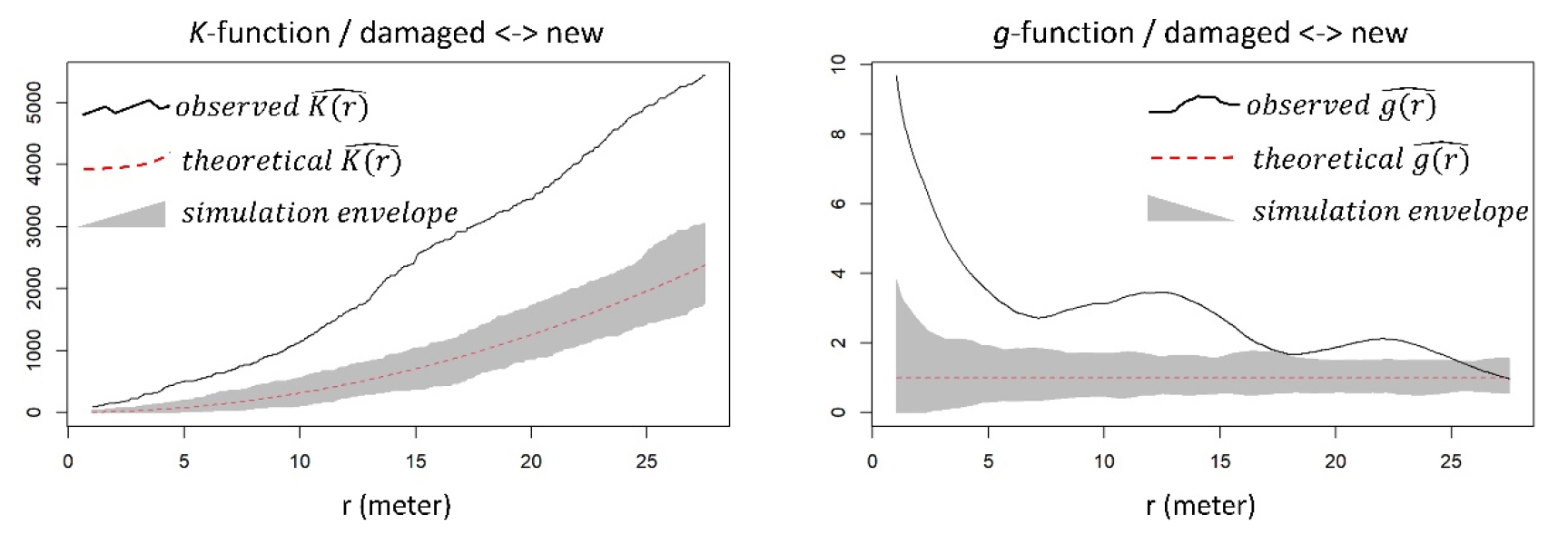
대홍수 이후 “새로 발생한 수목”의 공간적 분포를 이차 효과로 설명하고자, “피해를 받은 수목”과의 상대적 위치에 대한 분석을 진행하였다 (Fig. 6). 분석 결과, 사용한 분석 기법과 무관하게 모든 공간적 스케일에서 두 종류의 수목들이 서로 군집하는 것으로 파악되었다. 즉 패치 스케일에서도, 그리고 연구 지역 전체 스케일에서도 “새로 발생한 수목”은 2020년 대홍수가 발생하기 이전부터 존재하였던 기존 수목의 인근에 위치하는 경향을 보였다.
3.3 회복되지 않은 수목 (“dead”) 및 회복한 수목 (“recovered”)과 새로 발생한 수목 (“new”)의 공간적 관계
이전 절에서는 “회복되지 않은 수목”과 “회복한 수목”을 하나의 종류로 통합하여 “새로 발생한 수목”과의 관계를 규명하였다. 하지만 기존의 수목 중 회복 여부에 따라 “새로 발생한 수목”과의 공간적 관계가 상이할 수 있다. 이에 따라 본 절에서는 홍수 이후 “새로 발생한 수목”이 “회복되지 않은 수목”과 가지는 공간적 관계 (Fig. 7), 그리고 “새로 발생한 수목”이 “회복한 수목”과 가지는 공간적 관계를 구분하여 살펴보았다 (Fig. 8).
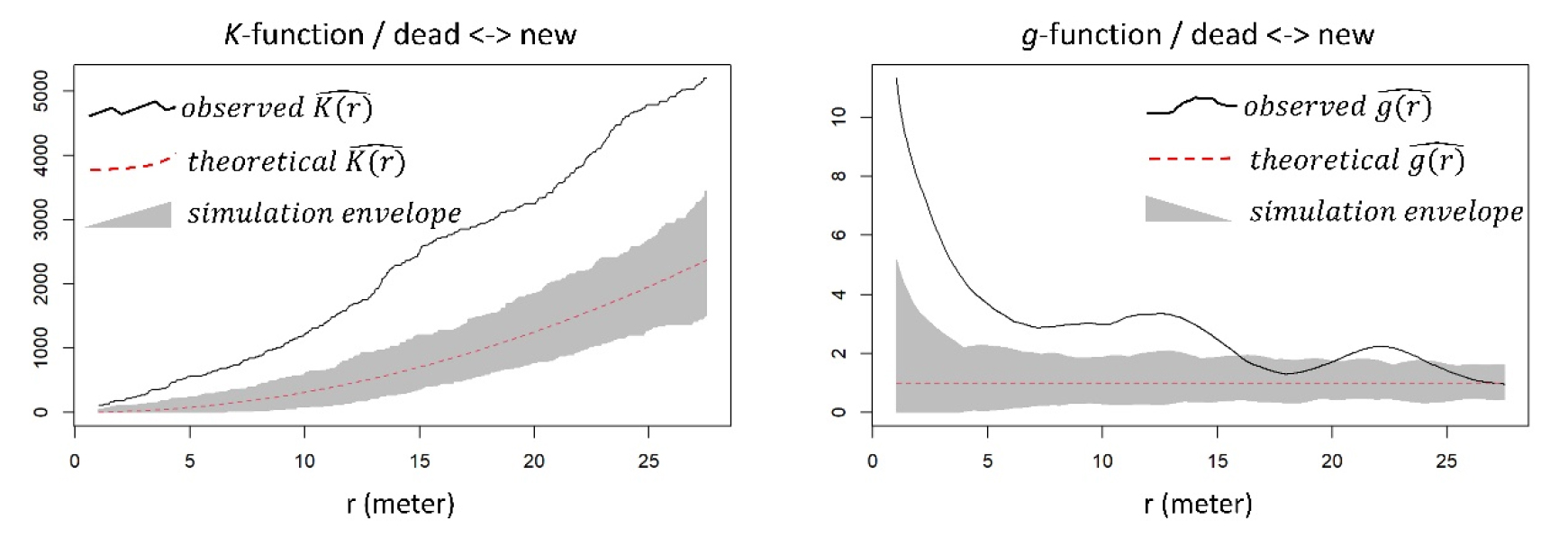
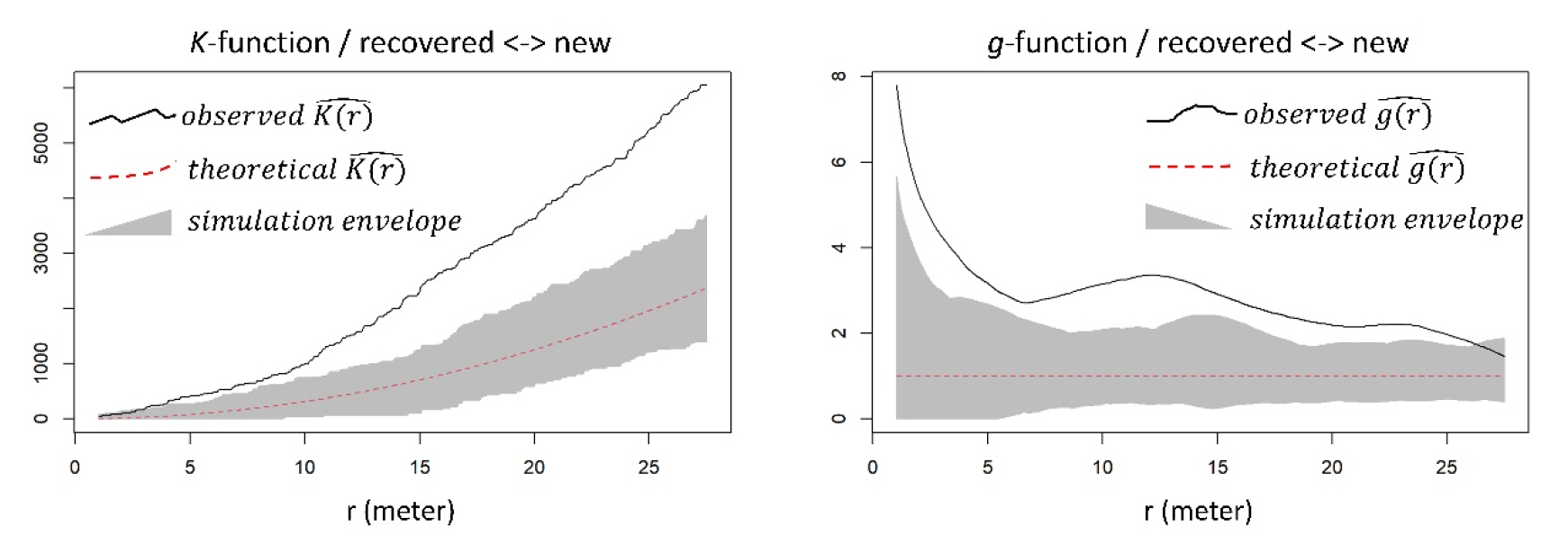
분석 결과, “새로 발생한 수목”은 대부분의 공간적 스케일에서 “회복되지 않은 수목” 및 “회복한 수목” 모두와 서로 군집하는 관계를 보였다. 다만 10 m 미만 반경의 좁은 스케일에 한정하여 정성적으로 비교한다면, 전자와는 서로 강하게 군집하는 반면, 후자와는 비교적 서로 약하게 군집하였다. Fig. 7에서는 함수 값 (observed K (r) and g (r))이 신뢰구간 (simulation envelope)보다 상당히 높은 곳에 위치하지만, Fig. 8에서는 함수 값이 신뢰구간보다 약간 높은 곳에 위치하고 있다. 즉 패치 스케일에서, “새로 발생한 수목”은 “회복한 수목”보다 “회복되지 않은 수목” 인근에 위치할 확률이 더 높았다.
4. 토 의
홍수 이후 새로 발생한 수목의 공간적 위치는 기존에 존재하던 수목의 공간 구조에 어느 정도 종속적이었다. 공간 점 분포 분석 결과, 패치 스케일에서도, 그리고 연구 지역 전체 스케일에서도 새로 생긴 수목은 홍수 이전부터 있던 수목의 위치와 통계적으로 유의미하게 인접하였다 (Fig. 6). 이를 통해 “새로 발생한 수목”이 연구 지역에 진입하는 과정에서 기존 수목으로부터 유기물을 공급받는 (Osterkamp and Hupp 2010) 등 긍정적 (facilitative) 효과를 받았을 것으로 추론할 수 있다. 하천 생태 복원 전략 중, 교란된 곳에 적응력이 높은 초기 천이 식생 (Salix spp.)을 인위적으로 식재하여 서식처 개선 효과 (habitat amelioration)를 통한 다른 식생의 진입을 유도하는 방법이 널리 사용된다 (González et al. 2015, Kauffman et al. 1997). 비록 본 연구지역에서 대홍수 이후 식생의 인위적인 도입이 이루어지지는 않았지만, 위와 같은 원리로 기존 수목이 새로 진입하는 수목의 성공적인 정착 확률을 증가시켰을 것이다.
앞서 언급한 대로 기존 수목과 새로 생긴 수목 간의 긍정적인 상호작용이 포착되었지만, 세부적으로는 그 반대의 관계 역시 작용하고 있다고 여겨진다. 공간 점 패턴 분석의 통계적 유의성 정도, 즉 군집하는 정도를 고려했을 때, 새로 생긴 수목은 완전히 죽은 수목보다 살아있는 수목과 더 약하게 서로 군집하였다 (Figs. 7 and 8). 이를 통해 촉진적 (facilitative) 상호작용 속에 가려진, 새로 생긴 살아 있는 수목과 기존의 살아 있는 수목 간의 경쟁적 (competitive) 관계를 미세하게나마 포착할 수 있었다 (see Tealdi et al. 2013). 하천 식생 간의 경쟁은 일사량 조절 (shading), 타감작용 (allelopathy), 근계경쟁 (root competition) 등을 통해 이루어진다 (Galindo et al. 2017). 하지만 현장 관찰 결과, 근계경쟁이 주요 프로세스일 것으로 예측될 뿐 구체적인 메커니즘을 밝히기 어려웠다.
대홍수 이전부터 존재하였던 수목이 교란을 겪으며 피해를 입었고, 일부 수목은 회복하지 못했지만 다른 일부는 회복하였다. 이러한 현상에 대한 이차 효과 분석 결과, 패치 내에서 개별 수목의 상대적인 위치로는 어떤 수목이 회복할지 여부를 예측할 수 없었다 (Fig. 4). 다만 피해를 입은 수목의 회복 여부는 수목이 위치한 고도에 의해 일부 설명된 바 있다 (Fig. 5). 이는 고도와 같은 환경 변수에 의해 수목의 회복 여부가 결정되는 일차 효과의 중요성을 보여준 결과라고 할 수 있으나, 본 연구에서는 교란 이후 회복하는 환경적 조건을 구체적으로 밝힐 수는 없었다는 한계가 있다. 대홍수의 맥락에서, 고도의 차이는 유속, 수심, 침수 시간 등의 차이를 야기한다. 그러므로 피해를 입은 수목이 회복되지 않은 이유가 유수에 의해 해당 개체에 큰 물리적 힘이 가해졌기 때문인지, 홍수 당시 해당 개체의 많은 부분이 물에 잠겼기 때문인지, 혹은 홍수 기간 동안 해당 개체가 전체적으로 오래 물에 잠겨 있었기 때문인지 규명할 수 없었다.
5. 결 론
대규모 홍수는 하천 생태계에 큰 교란으로 작용한다. 그 이후 생태계가 변화하는 과정을 연구함으로써, 기후변화의 맥락 속에서 하천을 효과적으로 관리하는 데 큰 도움이 될 수 있다. 하천 식생은 하도 (河道)를 변경하고 (Gurnell 2014, Gurnell et al. 2001), 홍수 시에 수위를 상승시키는 (Lee et al. 2021, Makaske et al. 2011) 등 하천의 수리적 맥락과 밀접한 관련을 맺고 있다. 더불어 하천 지형 변화 프로세스 역시 식생의 영향을 받기 때문에 (Corenblit et al. 2015a, 2015b, Kim 2021), 하천 식생에 대한 이해는 하천 생태계에 대한 통합적 접근에 필수적이다 (Woo et al. 2019). 홍수 후 수목의 회복과 새로운 수목의 발생을 설명하는 이차 효과를 규명함으로써, 강한 교란이 더 빈번하게 발생할 미래 사회에 새로운 하천 관리적 지식을 제공할 것으로 기대된다.
본 연구에서는 개별 수목의 상대적 위치를 통해 교란 이후 식생 군집의 변화를 설명하는데 주력하였지만, 일차 효과가 가지는 중요성을 과소평가할 수 없다. 환경 변수에 의해 식생의 분포가 결정된다는 시각과 수목 개체 간의 상호작용을 강조하는 시각은 모두 균형 있게 다루어져야 한다. 그러므로 고도를 통해 재생 여부를 설명한 본 연구의 결과를 출발점으로 하여, 향후 다양한 수리적 ․ 지형적 맥락에서 2020년 대규모 교란 이후 섬진강 하천 식생의 변화를 설명하려는 시도가 이어져야 할 것이다.
