

1. 서 론
2. 연구방법
2.1 흰수마자 포획 및 이주가능지 조사
2.2 인공종묘생산
2.2.1 친어 포획
2.2.2 산란유도, 부화 및 양성
2.2.3 유전다양성 분석
2.3 방류 및 모니터링
3. 결 과
3.1 이주 시도
3.2 인공종묘생산, 방류 및 모니터링
3.2.1 친어와 종묘생산된 치어의 유전구조 비교
3.2.2 치어방류지 선정
3.2.3 방류 및 모니터링
4. 고찰 및 결론
1. 서 론
환경부의 야생생물 보호 및 관리에 관한 법률에 근거한 멸종위기야생생물 I급 담수어류인 흰수마자 (Gobiobotia naktongensis)는 1 mm 이하인 모래가 90%이상을 구성하고 있는 서식지에 서식한다 (Chae 2004). 잉어과 (Cyprinidae) 모래무지아과 (Gobioninae)에 속하는 흰수마자는 전장 6 - 10 cm인 소형 어류로서 Mori (1935)가 낙동강의 영주와 함양에서 채집하여 신종으로 보고한 한국고유종이다 (Kim et al. 2014). 흰수마자는 수질이 양호하며 입자가 가는 모래여울이 잘 발달된 낙동강 유입 지류와 본류에서도 출현하며, 특히 낙동강 상류의 내성천, 중류의 감천과 황강 수계에서 비교적 많은 개체수가 출현하고 있는 것으로 보고되었다 (MLIT 2010).
한국의 대표적인 모래하천인 내성천은 모래하상이 넓게 펼쳐진 여울부가 발달되어 있는 하천 환경을 보유하고 있어 흰수마자의 최적 서식처로서 알려져 있다. 그러나 현재 영주다목적댐 건설공사가 이루어지고 있으며, 댐 건설 후 현재 유수역인 하천은 정수역으로 변하여 하천 여울에 주로 서식하는 흰수마자와 같은 유수성 어류가 타 지역으로 이동하거나, 댐에 의한 내성천 상하류 간 어류의 이동이 단절될 것으로 예상된다. 멸종위기 야생동식물을 관리하는 것은 고유 생물자원의 보전 및 생물다양성 확보와 함께 생태계를 유지하기 위하여 중요하며 (MLIT 2010, Ko et al. 2012), 특히 하천 공사, 골재 채취 등으로 인해 흰수마자의 서식처 파괴와 감소가 급격히 이루어지고 있기 때문에 본 종에 대한 보전 대책을 수립하는 것이 시급하다.
따라서 본 연구에서는 자연상태의 흰수마자 개체를 영주댐 수몰예정지인 내성천에서 포획하여 이들에게 적합한 하천 환경을 유지하고 있는 최적 서식지에 이주 방류하고, 축조되는 댐에 의한 생태통로 단절에 의하여 발생하는 흰수마자의 유전적 폐쇄을 방지하고 이들 집단의 유전적 동질성을 확보하기 위하여 내성천에서 포획한 흰수마자를 통해 인공종묘생산한 치어를 증강 방류하여 그 효과를 모니터링하였다.
2. 연구방법
2.1 흰수마자 포획 및 이주가능지 조사
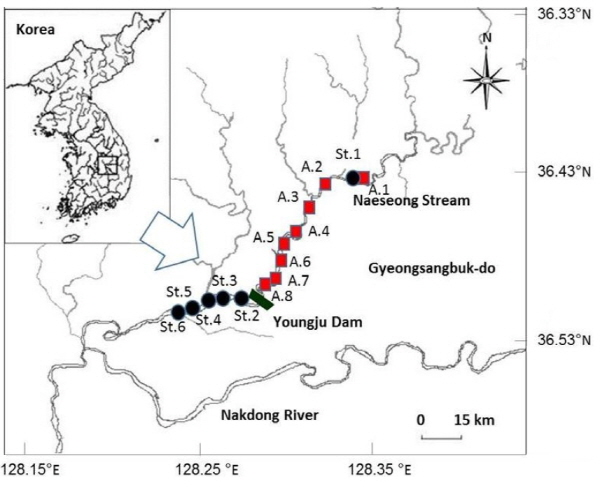
흰수마자의 포획 및 보관을 위하여 대구지방환경청의 포획․보관․훼손․방사허가 (2014-19호)를 받아 2014년 8월 7일 - 10월 31일 및 2015년 4월 1일 - 6월 30일까지 총 8차례에 걸쳐 영주댐 수몰예정지 내 8개 지점에서 흰수마자를 족대 (망목 5 mm × 5 mm)를 이용하여 포획을 시도하였다 (Fig. 1).
영주댐 수몰지 내에서 포획한 흰수마자의 적합 이주 가능지를 선정하기 위하여 2014년 6월 4일 영주댐 상류 St.1 및 하류 St.2-5의 5개 지점에서 무생물 및 생물 환경을 조사하였다 (Fig. 1). 하폭과 유폭은 디지털거리측정기 (Bushnell Sports 600, USA), 유속은 tachometer (Swoffer Model 2100, USA), 수심은 줄자를 이용하여 측정하였으며 하상구조는 Cummins (1962)의 기준에 따라 분류하였다. 흰수마자의 선호 미소서식처를 파악하기 위하여 각 조사지점에서 하상의 모래를 사각 채집통 (가로 40 cm × 세로 30 cm × 높이 20 cm)을 이용하여 채취하였다. 채집된 하상토는 0.106, 0.250, 0.500, 0.710, 1.00, 1.40, 2.00, 2.36, 2.80 및 3.35 mm 망목 크기의 체 (Retsch, AS 200)를 이용하여 입자 크기를 분석하였다.
|
Fig 1. A map showing the study sites for alternative habitat (St.1 - 5), optimal juvenile habitat (St.1 - 6) and sub-merged planned area of the dam construction (A.1 - 8). |
흰수마자의 먹이생물을 파악하기 위하여 조사지점의 흰수마자 개체군과 수서곤충 등을 직경 15 cm, 깊이 5 cm의 모래채집기를 이용하여 채집하였다. 채집된 생물 시료는 즉시 10% formalin에 고정하여 연구실로 이동한 후 해부현미경과 광학현미경상에서 Yoon (1995)과 Won et al. (2005)에 따라서 동정하였다. 이상의 조사 결과를 대표적인 흰수자마의 서식지 감천의 환경 조건 (Kim et al. 2014)과 비교하여 최적 이주가능지를 선정하였다.
2.2 인공종묘생산
2.2.1 친어 포획
흰수마자 인공종묘생산과 복원을 위한 포획 및 보관을 위하여 흰수마자의 산란기인 6-7월 중순 경에 맞추어 (Ko et al. 2013), 대구지방환경청의 포획․보관․훼손․허가 (2014-18호)를 받아 2014년 8월 9일 경북 예천군 호명면 담암리 및 용궁면 향석리 성저교 일대에서 족대 (망목 5 mm × 5 mm)를 이용하여 흰수마자 성어 40개체 (암컷 15, 수컷 25)를 포획하였다. 현장에서 암컷과 수컷을 마취시킨 후 복부와 생식공을 확인하여 친어로 적합한 개체를 선별하였다. 친어의 손상과 스트레스를 줄이기 위하여 50–100 ppm의 낮은 농도의 MS222로 마취 하였다. 선별된 친어는 산소 포장 후 실험실로 운반하였고 질병관리를 위하여 항생제와 염분 (5-7‰)에서 약욕을 실시하였다.
2.2.2 산란유도, 부화 및 양성
육안으로 복부와 생식공의 상태를 관찰하여 산란이 가능할 것으로 판단되는 암컷과 수컷을 선별하여 마취시킨 뒤 암컷은 Ovaprim 0.5 ml/kg, 수컷은 HCG 40 IU/g의 농도로 복강에 주사하였다. 암컷과 수컷을 1:2 또는 1:3의 비율로서 광조건, 암:명 = 10:14, 수온 25°C, 용존산소 8 mg/L 이상의 조건이 유지되는 100 L 수조에 수용하였다. 암막을 이용하여 완전하게 빛을 차단하여 산란이 용이하도록 하였으며, 수용 10 - 12시간 후에 산란이 이루어졌다.
흰수마자의 수정란은 산란수조 내에서 그대로 유지하여 부화할 때까지 사육하였다. 흰수마자 친어는 산란 후 어란을 섭식하는 행동을 보이기 때문에 산란 이후 다른 수조로 옮겼다. 부화는 수정 후 18-20시간 사이에 이루어졌다. 부화된 자어가 최소한의 영향을 받도록 매우 약하게 공기 기폭만 시켜주었으며, 죽은 자어는 스포이드를 이용하여 제거하였다. 3-4일 이후 초기먹이를 섭식하기 시작하면 자어보다 큰 사이폰을 이용하여 5 L 박스로 옮겨서 관리하였다. 초기먹이로는 5-7일까지는 물벼룩을 공급하였으며, 이후 60일까지는 Artemia를 공급하였다. 먹이는 1일에 3회 공급하고 바닥에 먹고 남은 먹이찌꺼기는 스포이드로 제거하였으며, 매일 1/2씩 사육수를 환수하였다. 100일 경과 후부터는 215 L 이상의 순환식 여과수조로 옮겨 유수를 서서히 흘려 보내주고 실지렁이를 매우 가늘게 잘라 먹이로서 공급하였으며, 먹이 공급 이후 남은 찌꺼기는 지속적으로 제거하였다.
먹이공급에 있어 순화하는 과정에서 질병에 의한 폐사개체가 발생할 수 있으므로 사육수는 초기 염분 3‰을 유지하여 관리하고, 질병 발생시 OTC 처리 및 염분을 7-8‰까지 상승시켜 약욕한 후 사육수의 80%를 환수하여 대량폐사가 일어나지 않도록 관리하였다. 또한 먹이로 공급하는 실지렁이는 소독제를 이용하여 소독하였으며, 2-3일 간 유수를 흘려주고 지속적으로 세척을 하여 질병원의 유입을 최소화시켜 먹이로 공급하였다. 종묘생산한 흰수마자 치어와 자연산 치어의 구분을 위하여 종묘생산 치어에 VIE 형광표지를 삽입하였다.
2.2.3 유전다양성 분석
유전다양성 분석을 위해 현재 개발되어 있는 흰수마자와 돌상어의 microsatellite 마커를 이용하였으며 (MOE 2011), 각 마커의 증폭은 PCR PreMix (Bioneer, Korea)를 이용하여 50-100 ng의 주형 DNA와 0.5 pmole의 primer를 포함한 총 부피 20 μL에서 실시하였다. PCR 조건은 처음 95°C에서 15분간 DNA를 변성한 다음 95°C에서 20초간 변성, 58°C에서 40초간 annealing, 72°C에서 1분간 extension을 35회 반복한 후 최종 DNA 합성을 72°C에서 15분간 하였다. 증폭된 산물은 1.5% agarose gel 상에서 전기영동으로 일일이 결과를 확인하여 희석 배율을 결정하였다. 희석한 PCR 산물과 Genscan™ 400HD (ROX) size standard, HiDimixture를 섞고 95°C에서 5분간 변성시킨 후 AVI 3730xl DNA analyzer (Applied Bio-systems, USA)를 이용하여 유전자형을 분석하였다. 대립유전자들의 정확한 크기는 GenescanTM 400HD (ROX) size standard를 이용하여 Peak ScannerTM Softwarever.1.0 (Applied Biosystems, USA)으로 분석하였다. 대립유전자의 크기가 결정된 데이터는 null allele, large allele dropout, scoring errors 유무와 분석 시 발생할 수 있는 입력오류를 판별하기 위해 집단별로 나타난 genotype을 MICRO-CHECKER 2.2.3를 이용하여 검사하였다 (Van Oosterhout et al. 2004). 기술 통계적 수치 (대립유전자 수, number of allele per locus; 기대치/관찰치 이형접합률, expected/observed heterozygosity; 다형성 정보 지수, polymorphism information content PIC; 대립유전자 빈도, allele frequency)는 Cervus 프로그램 ver. 3.0 (Marshall et al. 1998)을 이용하였으며, 증폭 안 된 개체가 존재함에 따라 allelic richness 수치는 FSTAT 프로그램 ver. 1.2 (Goudet 1995)를 이용하여 산출하였다.
2.3 방류 및 모니터링
인공종묘생산한 흰수마자 치어의 적합방류지를 선정하기 위하여 2014년 9월 20일 - 21일 동안 영주댐 상류 St.1과 하류 St.2 – 6의 총 6개소를 대상으로 이주 적합지와 동일한 방법으로 무생물 및 생물 환경을 조사하였다.
인공종묘생산한 흰수마자 치어 총 5,000개체를 2014년 10월, 2015년 5월 등 2차례에 걸쳐 방류 후에 모니터링을 실시하였으며, 모니터링 기간은 방류 직후부터 영주댐 준공시까지로 설정하였다. 모니터링을 위하여 족대 (망목 5 mm × 5 mm)를 이용하여 포획하여 방류지로부터 상하류 이동양상과 재포획률 (방류개체의 생존율 추정)을 분석하였다. 미소분산은 방류지점 중심 상하류 1 km, 확대분산은 방류지점 상하류 10 km (2 km 간격)를 대상으로 총 11지점에서 조사하였다.
3. 결 과
3.1 이주 시도
영주댐 수몰지에서 포획한 흰수마자의 최적 이주가능지를 선정하기 위하여 영주댐 상하류의 St.1 - 5의 총 5개 후보지에서 하천 환경을 조사하여 감천의 자연 서식지 (Kim et al. 2014)와 비교한 결과, 이들 후보지의 물리적 환경은 모두 양호한 것으로 판단되었다. 특히 하상 입도 조성은 감천의 자연 서식 수역에서 1 mm 이하의 모래가 80% 이상을 차지하였는데, 조사지점 St.4가 78.1%, St.5가 79.7%로서 이와 가장 유사하였다. 흰수마자의 주요 먹이인 깔다구과 유충의 개체수 St.4와 St. 5가 다른 지점에 비해 높은 것으로 확인되었다. 한편 2014년 6월 조사시 흰수마자는 St.5 (미호교)에서만 6개체를 확인할 수 있었다. 이상을 종합하면 흰수마자의 최적 이주지로서 St.4 (오신교)와 St.5 (미호교)가 적합할 것으로 판단되었다.
이주개체를 확보하기 위하여 영주댐 수몰 예정지인 댐 상류 내성천 20 km까지의 A.1 – 8 지점에서 2014년 7월 - 2015년 7월까지 총 8차에 걸쳐 개체 조사를 시행하였으나 흰수마자를 채집하지 못하였다. 따라서 이주가능지로의 이주도 이루어지지 못하였다.
3.2 인공종묘생산, 방류 및 모니터링
3.2.1 친어와 종묘생산된 치어의 유전구조 비교
내성천에서 포획한 흰수마자 친어 40개체를 통해 2014년 8월 - 10월 동안 5,000개체의 치어를 인공종묘생산하였다. 친어와 치어 집단 간의 유전 다양성을 비교하기 위하여 흰수마자 친어 27개체와 생산된 치어 64개체간의 유전구조를 10개 microsatellite 마커를 이용하여 비교하였다. 각 집단별, 마커별 대립유전자수 (number of alleles, Na), 동일샘플수 상의 대립유전자 수를 나타내는 대립유전자 풍부도 (allelic richness, A), 이형접합률 관찰치 (observed hetero-zygosity, Ho), 이형접합률 기대치 (expected hetero-zygosity, He), 그리고 다형성 정보지수 (polymorphic information content, PCI)를 Table 1에 나타내었다. 친어 집단과 종묘생산된 치어 집단간의 마커별 대립유전자 빈도 (allele frequency) 분석 결과 생산된 치어의 대립유전자 수는 친어 집단과 비슷한 값을 보였다. 또한 분석에 이용한 10개의 마커별 주요 유전자형의 대립유전자는 변하지 않았으며, 두 집단간에 빈도차만 있었다. 이는 치어를 인공종묘생산하기 위하여 이용한 친어의 개체 선별이 성공적이었음을 의미하였다.
친어 집단의 평균 대립유전자 수는 16개이었으며, 생산된 치어의 평균 대립유전자 수는 14.8개로 생산된 치어 집단의 대립유전자수가 약간 적게 나타났다 (Table 1). 친어 집단의 대립유전자 풍부도는 14.4개였으며, 생산된 치어 집단은 13.1개로서 치어 집단의 대립유전자 풍부도가 약간 적었다. 친어 집단과 치어 집단의 평균 이형접합률 관찰치 (Ho), 이형접합률 기대치 (He) 및 다형성 정보지수 (PCI)는 서로 유사하였다. 따라서 인공종묘생산한 치어 집단과 친어 (자연) 집단 간의 유전적 차이는 거의 없는 것으로 확인되었다.
이상의 결과를 종합하면 인공종묘생산에 사용된 어미의 수가 치어의 유전 다양성을 감소시킬 정도로 적지 않았으며, 친어의 유전 다양성이 종묘생산된 치어 집단으로 잘 전달된 것을 의미한다. 따라서 본 연구에서 생산한 치어를 자연에 방류했을 시 자연집단의 유전다양성 감소 또는 유전자 교란 등의 문제를 야기하지 않을 것으로 판단되었다.
3.2.2 치어방류지 선정
인공종묘생산한 흰수마자의 최적 방류지 선정을 위하여 영주댐 상하류의 St.1 - 6까지 총 6개 후보지에 대하여 이주가능지와 동일한 기준으로 평가하였다. 이주가능지 선정과 마찬가지로 감천의 자연 서식지 (Kim et al. 2014)와 환경을 비교하면 이곳의 물리적 환경은 모든 지점이 양호하였다 (Table 2). 특히 흰수마자 당년생 치어가 서식하는 수역은 1 mm 이하의 모래가 85% 이상을 차지하며 (Kim et al. 2014), St.5 지점 하상의 1 mm 이하의 모래 조성이 98.7%로서 다른 지점보다 풍부하였다 (Table 3). 또한 흰수마자의 주요 먹이인 깔다구과 유충의 개체수 및 습중량 비교시 St.3와 St.5가 다른 지점보다 많았다 (Table 4). 따라서 St.5 (미호교) 지점이 치어 방류지로서 가장 적합한 것으로 판단되었다.
3.2.3 방류 및 모니터링
종묘생산한 흰수마자 2,000개체의 1차 방류는 2014년 10월 15일에 사전 선정한 St.5와 6 (경상북도 예천군 보문면 미호리 미호교 인근) 지점에서 이루어졌다. 방류 후 1, 3, 7, 15 및 30일차에 개체수 조사를 실시하였다. 방류 후 1일과 3일에 미소분산 모니터링을 위하여 방류지점으로부터 상류 50 m와 하류 150 m 구간까지 분산 상태를 조사한 결과 각각 43개체와 4개체를 확인하였다. 그러나 방류지 상하류 10 km 구간에서 확대분산이 이루어지는 방류 후 7일 이후부터는 모든 지점에서 방류개체를 확인할 수 없었다. 이러한 이유는 이 시기에 내성천 유역에서 집중호우가 발생하여 내성천의 수위가 상승하였고 유량과 유속이 급속히 증가하여 하상이 교란되었기 때문일 것으로 생각된다. 1차 방류일로부터 6개월차 (2015년 4월 16-17일), 7개월차 (2015년 5월 13-14일), 8개월차 (2015년 6월 26-27일) 및 9개월차 (2015년 7월 6-7일)에 실시한 모니터링에서 모든 지점에서 1차 방류 개체는 확인되지 않았다. 다만 조사기간 중 일부 지점에서 2, 3년생 성어가 확인되었다. 2차 방류 치어 모니터링 과정 중 2015년 5월 28-29일 St.3에서 1차 방류개체 1개체가 포획되었으며, 이 개체는 암컷으로서 다소 복부가 팽대되어 있는 것으로 보아 정상적으로 생식과정이 진행 중인 것으로 확인되었다.
종묘생산한 흰수마자 치어 3,000개체의 2차 방류 역시 1차와 마찬가지로 St.5 지점에서 2015년 5월 13일 이루어졌으며, 방류 후 1, 3, 7, 15, 30일차 개체수 조사를 실시하였다. 미소분산이 이루어지는 방류 1일차 및 3일차에는 1차 방류 때와 마찬가지로 방류지점으로부터 상류 350 m 및 하류 240 m 구간까지 각각 42개체 및 29개체가 확인되었다. 1차 방류 때와는 다르게 확대분산이 이루어지는 방류 후 7일차부터 30일차 (2015년 6월 15-16일)까지 St.6과 7 지점에서 일부 개체가 확인되었으며, 2개월차가 되는 시점인 2015년 7월 6-7일에는 모든 지점에서 방류 개체는 확인되지 않았다.
4. 고찰 및 결론
수몰이 예상되는 영주댐 상류 내성천 구간에서 흰수마자를 포획하기 위하여 2014년 7월부터 2015년 7월까지 8차례에 걸쳐 개체수 조사 및 포획을 시도하였으나 서식이 확인되지 않아 기 선정한 St.5 지점으로 이주는 이루어지지 않았다. 이는 최초 수몰지 내 포획이 시작된 2014년 7월 이전에 서식 가능한 흰수마자 개체들은 산란기인 7 - 8월에 맞추어 영주댐 건설지역 하류인 내성천 최하지점 또는 인접 낙동강 본류 수역으로 산란을 위해 이동하여 (Kim et al. 2014) 산란을 마친 후에 댐 상류 수역으로는 이동하지 않은 것으로 추정되나, 향후 지속적인 모니터링을 통한 이에 대한 추가 조사가 필요하다고 생각된다.
금강과 낙동강의 흰수마자 집단의 유전자다양성 분석 결과 수계간 유전적 거리, 유전적 분화도 및 변이가 뚜렷이 구분되었기 때문에 인공종묘생산된 치어는 사용된 친어에 따라 수계를 명확히 구분하여 방류하여야 하며 (MLIT 2010), 이에 따라 본 연구에서도 내성천에서 포획한 친어를 통해 종묘생산한 치어를 내성천에 방류하여 유전적 동질성을 확보하고자 하였다. 종묘생산한 흰수마자 2,000개체의 1차 방류 모니터링을 위하여 방류지 상하류 10 km 범위 내 11개의 조사지점을 주기적으로 방문하여 흰수마자 개체군을 조사한 결과, 방류 직후인 1, 3일차에는 방류지를 중심으로 각각 43개체, 4개체가 포획되었으나 7일차인 2014년 10월 21일 이후부터는 방류지역을 중심으로 조사범위 내에서 발견되지 않았다. 이는 2014년 10월 이후 많은 양의 가을비로 인해 내성천의 수위가 급격히 상승되고 유속이 증가하면서 하상 교란이 발생하여 (2014년 10월 21일 일강우량 영주 101 mm, 봉화 59 mm) 방류한 치어들이 보다 신속히 분산된 것으로 추정된다. 1차 치어방류 시점으로부터 6개월차인 2015년 4월 조사시에도 전체 조사지점에서 포획된 흰수마자는 모두 2, 3년생 성어로서 인공종묘생산한 흰수마자는 발견되지 않았다. 특히 2015년 5월 2차 방류치어 조사시 1차 방류치어가 방류지점 6 km 상류에서 1개체 포획되었고, 이 개체는 암컷으로서 다소 복부가 팽대되어 있는 것으로 보아 정상적으로 성분화가 진행 중으로 확인되었으므로 산란연령인 만 2년생에 도달하는 2016년에는 번식에 참여할 가능성이 높을 것으로 판단되었다. 이는 인공종묘생산한 흰수마자가 내성천의 자연생태환경에 적응하고 있다는 가능성을 제시하는 것으로 향후 지속적인 연구를 통한 검토가 필요하다.
종묘생산한 흰수마자 3,000개체는 2차로 2015년 5월 13일에 St.5와 인근 지역을 중심으로 방류하였으며, 1차 방류와 동일한 방법으로 흰수마자 개체군을 모니터링하였다. 방류 후 1, 3일차에는 방류지를 중심으로 30 - 40개체가 포획되었으나 7일차, 15일차 및 30일차인 2015년 5 - 6월까지 방류지점 및 상하류 10 km 범위 내에서 2-3개체만 발견되어 방류 후 시간이 지남에 따라 포획 개체수가 감소하였다. 5-6월 포획된 2차 방류치어 3개체는 방류개체 중 비교적 크기가 큰 개체들이었으며, 모두 복부 풍만도가 높은 것으로 보아 자연 먹이를 충분히 섭식하는 것으로 확인되었다. 방류 2개월차인 2015년 7월 6 - 7일에는 모든 지점에서 방류개체는 확인되지 않았다. 2차 방류의 경우 봄철 수위변동이 미비해 하상 안정성이 높은 시기인 5월에 방류하여 방류 치어의 자연 적응도와 분산효과에 영향을 미친 것으로 파악되며, 현재 모니터링 지점인 20 km 구간에서 자연산 만 1년생 치어는 발견되지 않고 본 사업으로 방류된 치어만 확인되고 있는 점으로 보아 내성천 종묘생산 흰수마자의 방류 및 확산이 지속적으로 진행되고 있는 것으로 판단된다. 향후 주기적이고 지속적인 모니터링을 통한 본 종묘생산 및 복원 효과의 장기적인 관찰과 평가가 필요할 것으로 보인다.
