1. 서 론
온실가스 배출량 증가로 인한 지구적 기후변화로 기후대가 고위도로 이동하고 식물 생육기간이 2일/10년이 증가하였다 (IPCC 2021). 한반도에서도 1912년부터 2020년까지 연평균기온이 0.2°C/10년 상승하였고 연강수량이 17.7 mm/10년 증가하였다 (KMA 2021). 이러한 기후변화에 따라 전지구적 생물종 및 식생분포대가 변화하고 있으며 (WWF 2015), 특히 한국의 난대식생대가 확장되고 국내 분포하지 않는 난아열대 식물이 점차 유입, 정착할 수 있을 것으로 예측되고 있다 (NIFS 2021, NIFS 2022).
기후변화에 따른 난대식생대와 난아열대 식물종의 분포 변화를 예측하기 위하여 일반적으로 일반화 가법 모형, Maxent 등의 다양한 종 분포 모형이 활용되고 있다 (Cho and Lee 2015, NIFS 2021, NIFS 2022). 이러한 종 분포 모형은 한 생물종에 대하여 특화된 예측 모형을 만들기는 쉽지만 여러 종에 동시에 적용할 수 있는 일반적인 모형을 만들기 적합하지 않을 뿐만 아니라 구축된 모형을 다른 목적으로 재이용하기 하기가 어렵다 (Elith and Leathwick 2009, Santini et al. 2021). 최근 종 분포 모형의 한계를 보완하기 위하여 다변량 환경 유사도 표면 (multivariate environmental similarity surfaces, MESS)을 통한 유사도 방법이 활용되고 있다 (Elith et al. 2010). MESS는 다차원 공간에서 두 집단의 변수들 사이에 얼마나 유사한지 나타내는 지표로서 음수와 양수를 모두 포함하는 그 값이 클수록 두 지역의 환경이 유사하다는 것을 나타낸다 (Elith et al. 2010). 이 분석법은 유사/비유사의 기준이 명확하고, 전체 공간 분포 정보를 동시에 고려하기 때문에 유사도를 추정하는데 강점이 있으며, 많은 대상 종에 대하여 간단한 모형으로서 적용할 수 있는 범용성이 큰 장점을 가지고 있다 (Owens et al. 2013, Mesgaran et al. 2014). 따라서 기후변화에 따라서 다수의 동아시아 난대 식물종이 한국에 분포할 가능성을 평가하는데 MESS가 유용할 것으로 판단된다.
본 연구의 목적은 동아시아 중국와 일본에 분포하지만 한국에 유입되지 않은 난대 목본식물이 한반도에 잠재적으로 분포할 가능성을 평가하기 위하여, 1) 기후 요인 중 난대 목본식물종의 분포 결정 요인을 추출하고, 2) MESS를 이용하여 동아시아의 난대 목본식물종의 전지구적 분포지와 한국 난대식생대의 기후변수 유사도를 추정하며, 3) 이를 종합하여 동아시아 난대 목본식물종의 한국의 잠재분포 가능성을 평가하는 체제를 구축하고자 하였다.
2. 연구 방법
한국에서는 분포하지 않고 있으나 지리적으로 가까워 이입 가능성이 높은 난대 목본식물종을 동아시아인 중국과 일본에서 수집하여 목록을 구축하였다. 중국의 난대 식물종은 Wang et al. (2018)에서 난대 식물로 구분되는 종을, 일본의 난대 식물종은 NIFS (2012)의 난대 식물 목록에서 수집하였다. 이들 목본식물 중에서 한국에 분포하거나 식재된 식물종은 제외하였다. 수집된 종은 WFO (2022)에 따라서 분류학적 실체를 검증하고 학명을 정리하였다. 수집된 난대 목본식물종의 분포 정보는 GBIF (2022)에서 점 분포 자료로 수집하였다. 수집할 때 식재 등에 의한 인위적 분포와 과거 화석 표본 자료는 제외하였다.
본 연구에서는 동아시아 난대 목본식물종이 한국에 분포할 수 있는 가능성을 평가할 수 있는 요인으로서 기후만을 고려하였다. 따라서 기후요인에 의하여 이들 식물이 한반도에서 먼저 정착할 수 있는 지역을 난대 식생기후대로 가정하였다. 난대식생대는 한랭지수 (coldness index)를 계산하여 한랭지수가 -10°C보다 큰 지역으로 정의하였다 (Kira 1945, Yim and Kira 1975, Kira 1991). 한랭지수는 Worldclim (2022)에서 2.5' × 2.5' 해상도로서 현재 월평균기온 자료를 활용하여 QGIS.org (2022)의 격자 계산기에서 계산하였다.
난대 목본식물종의 분포에 영향을 미치는 기후 자료는 Worldclim (2022)에서 제공하는 19개의 생물기후 자료를 이용하였다. 이 기후 자료를 한국의 난대식생대 격자와 선정된 난대식물종의 한반도를 제외한 전지구적 분포 격자에서 추출하였다. 이들 생물기후 변수 간 관계 및 한반도 난대식생대와 난대식물종별 분포지의 관계를 파악하기 위하여 R 환경 (R Core Team 2022)에서 ‘vegan’ 패키지의 ‘rda’ 함수를 이용하여 주성분 분석 (principal component analysis, PCA)를 실시하였다 (Oksanen et al. 2022).
중국과 일본에 분포하지만 아직 국내에 분포하지 않는 난대 목본식물의 한국에 잠재분포 가능성을 평가하기 위하여, 이들 식물종의 분포에 영향을 미치는 주요 기후 변수를 선별한 후 다변량 회귀분석을 R 환경 (R Core Team 2022)에서 수행하였다. 회귀 분석에서 기후 변수의 다중공선성 (multicollinearity)을 해소하기 의하여 ‘cor.test’ 함수를 이용하여 변수간 Pearson 상관분석을 실시하였다. 상관분석 결과에서 상관계수가 0.75 이상이거나 -0.75 이하인 기후변수를 상관계수의 절대값 총합이 가장 큰 변수만 남기고 제거하였다 (Padalia et al. 2014, Negrini et al. 2020). 한국의 난대 식생대 기후 환경과 동아시아 난대 식물 분포지의 기후 환경 사이의 유사도를 추정하기 위하여 다변량 환경 유사도 표면 (multivariate environmental similarity surfaces, MESS) 분석을 수행하였다. 한국 난대식생대 격자와 동아시아 난대 식물 종별 본포 격자에서 선정된 기후변수 사이에 MESS를 ‘modEvA’ 패키지의 ‘MESS’ 함수를 이용하여 계산하였다 (Barbosa et al. 2013). MESS로 산출된 환경 유사도 지수는 양의 값으로 클수록 두 지역의 환경이 더욱 유사하며, 음의 값으로 적을수록 환경이 더욱 큰 차이가 있으며, 0이면 최소로 환경 범위가 겹친다는 것을 의미한다 (Elith et al. 2010). 동아시아 난대 목본식물종의 한국 난대식생대에서의 잠재분포 가능성을 평가하는 체제를 구축하기 위하여 MESS 환경 유사도 지수를 종속변수로 하고 선정된 기후 변수를 독립변수로 하여 ‘lm’ 함수 및 ‘step’ 함수를 이용하여 단계적 변수선택 회귀분석을 실시하였다. 이때 모형에서 독립변수의 상대적 중요도는 ‘asbio’ 패키지의 ‘partial.R2’ 함수를 이용하여 부분 R2로 추정하였다 (Aho 2022). 최종적으로 각 난대 목본식물종의 분포지에서 기후 변수 평균 값을 모형에 입력하여 환경 유사도를 산출하여 잠재분석 가능성을 평가하였다.
3. 결과 및 고찰
현재 한국에서는 분포하지 않지만 우리나라에 이입되어 생육이 가능할 수 있는 동아시아의 난대 목본식물종을 수집한 결과, 중국 342 종과 일본 49 종이었고 두 국가에서 공통으로 나타나는 종이 7 종으로서 연구대상 후보종이 총 384 종이었다. GBIF (2022)에서 수집한 이들 후보종의 분포지는 총 84,549 개소이었다. 분포지 수가 10 개보다 적은 65 종은 신뢰성 확보가 어렵다고 판단하여 연구대상에서 제외하여 총 319 종을 본 연구에 이용하였다 (Appendix 1). 선정된 동아시아 난대 목본식물종의 분포지는 아시아에서는 적도를 포함한 저위도 지역에 특히 남반구에서 위도 높은 지역까지 분포하였다 (Fig. 1). 아프리카와 남미에서는 내륙 열대기후대보다는 해안을 따라서 다소 위도가 높은 곳에 분포하였다. 북미와 유럽에서는 아시아보다는 위도가 높은 곳에 분포하고 있었다. 한편 한랭지수가 -10°C보다 큰 난대식생대는 제주 해안 그리고 한반도의 남쪽에 특히 해안을 따라서는 위도가 보다 높은 곳에 위치하였다 (Fig. 1).
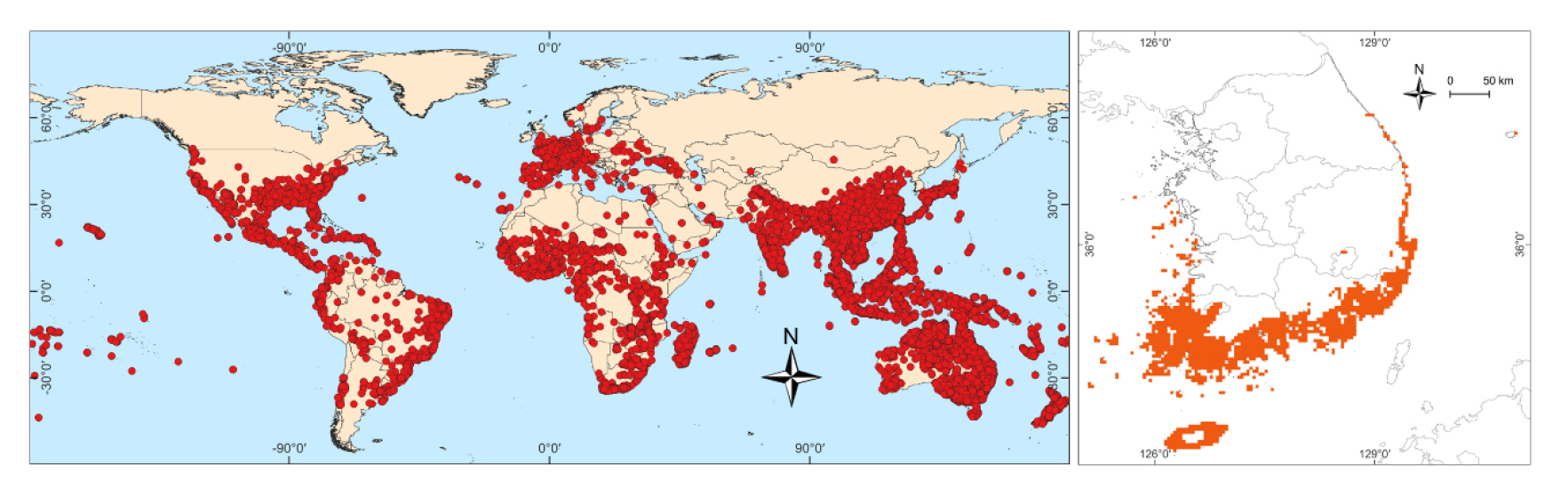
Fig. 1
A map showing the global distribution of warm-temperate woody plants distributed in China and Japan not in Korea (left) and the warm temperate vegetation zone of Korea according to the coldness index (Kira 1991) (right).
선정된 동아시아 난대 목본식물종 분포지와 한국 난대식생대의 19개 생물기후 변수를 이용한 PCA에서 1축과 2축이 전체변이의 각각 39.7%와 26.8%를 차지하였다 (Fig. 2). 장소의 배열에서 한국 난대식생대 지소가 동아시아 난대 목본식물종 분포지의 왼쪽에 좁게 포함되어 있었다. 한편 기후변수의 배열에서 대부분의 온도 관련 변수 (T01 – T11)은 제1축의 양끝에, 강수량 관련 변수 (P01 – P08)은 제2축 양끝에 위치하였다. 따라서 온도 관련 변수 집단과 강수량 관련 변수 집단이 각각 서로 강하게 관련되어 있다는 것을 알 수 있었다. 이에 따라서 기후변수의 다중공선성을 해소하기 위한 상관분석에서 T11 (평균온도일교차)을 제외한 모든 온도 관련 변수 (T01 – T10)가 높은 상호 상관을 가졌다 (Table 1의 황색 음영). 또한 강수량 관련 모든 변수 (P01 – P08)가 서로 깊은 상관을 가졌다 (Table 1의 청색 음영). 상호 상관관계를 갖는 변수군에서 다른 변수와의 상관계수 합이 가장 큰 변수를 대표 변수로 선정하였는데, T01 – T10 변수군에서는 T07 (최한 사분기 평균기온)을 P01 – P08 변수군에서는 P04 (연강수량)이 선택하였다.
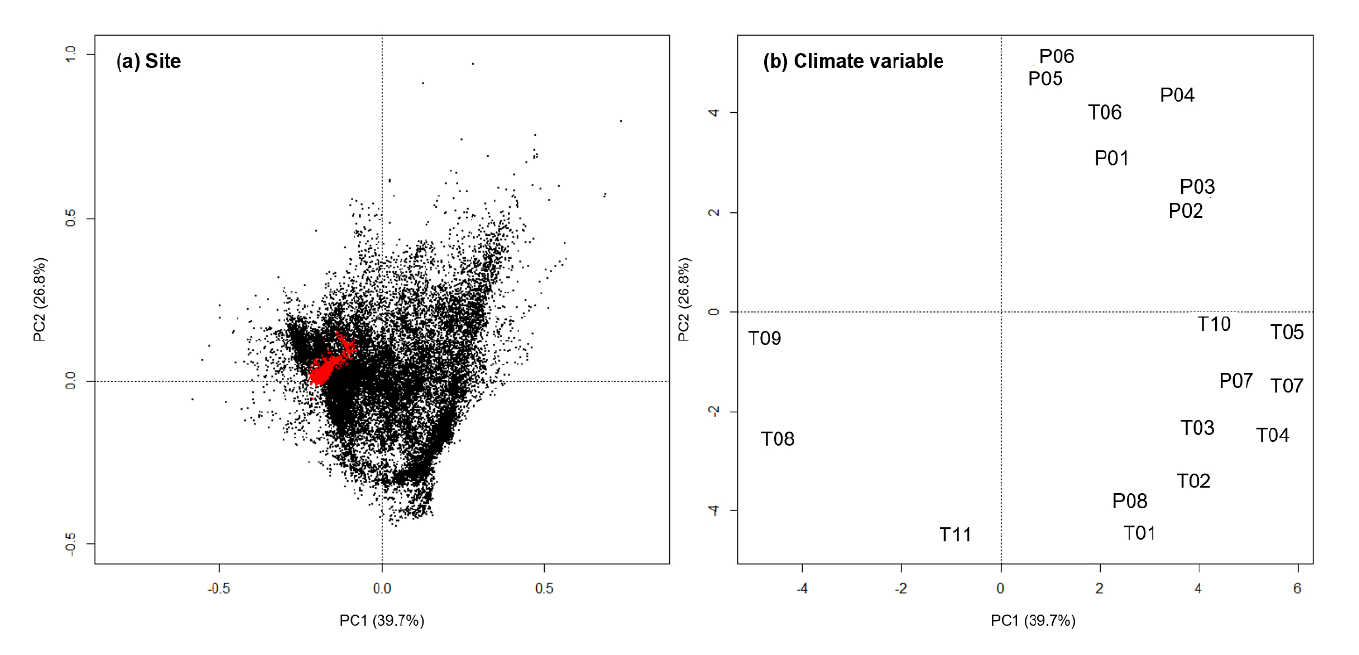
Fig. 2
Biplots of principal components analysis (PCA) of climates in the distribution sites of warm-temperate woody plants of East Asia (black dot) and in the warm-temperate vegetation zone of Korea (red dot) (T01 = max temperature of warmest month, T02 = mean temperature of warmest quarter, T03 = mean temperature of wettest quarter, T04 = annual mean temperature, T05 = min temperature of coldest month, T06 = mean temperature of driest quarter, T07 = mean temperature of coldest quarter, T08 = temperature annual range, T09 = temperature seasonality, T10 = temperature isothermality, T11 = mean temperature diurnal range, P01 = precipitation of warmest quarter, P02 = precipitation of wettest month, P03 = precipitation of wettest quarter, P04 = annual precipitation, P05 = precipitation of driest month, P06 = precipitation of driest quarter, P07 = precipitation of coldest quarter, P08 = precipitation seasonality.
Table 1.
Pearson's correlation coefficients between climate variables. Italic numerals indicate the significance at α = 0.01 and bold numerals indicate values greater than 0.75 as an absolute value. Shading of the same color indicates groups of variables that are strongly correlated with each other. Abbreviations refer to Fig. 2
|
T01
|
T02
|
T03
|
T04
|
T05
|
T06
|
T07
|
T08
|
T09
|
T10
|
T11
|
P01
|
P02
|
P03
|
P04
|
P05
|
P06
|
P07
|
P08
|
|
T01
| 1 | | | | | | | | | | | | | | | | | | |
|
T02
| 0.94 | 1 | | | | | | | | | | | | | | | | | |
|
T03
| 0.75 | 0.83 | 1 | | | | | | | | | | | | | | | | |
|
T04
| 0.77 | 0.86 | 0.79 | 1 | | | | | | | | | | | | | | | |
|
T05
|
0.44
|
0.58
| 0.75 | 0.89 | 1 | | | | | | | | | | | | | | |
|
T06
|
0.50
|
0.52
|
0.31
| 0.78 | 0.84 | 1 | | | | | | | | | | | | | |
|
T07
|
0.57
|
0.67
|
0.65
| 0.95 | 0.97 | 0.85 | 1 | | | | | | | | | | | | |
|
T08
|
0.19
|
0.00
|
-0.15
|
-0.46
| -0.8 | -0.75 | -0.75 | 1 | | | | | | | | | | | |
|
T09
|
-0.06
|
-0.15
|
-0.25
|
-0.63
| -0.86 | -0.75 | -0.83 | 0.90 | 1 | | | | | | | | | | |
|
T10
|
0.18
|
0.19
|
0.27
|
0.60
| 0.76 | 0.75 | 0.77 | -0.76 | -0.88 | 1 | | | | | | | | | |
|
T11
|
0.50
|
0.22
|
0.10
|
0.15
|
-0.15
|
0.12
|
0.07
|
0.49
|
0.08
|
0.19
| 1 | | | | | | | | |
|
P01
|
-0.17
|
0.07
|
0.32
|
0.12
|
0.18
|
-0.11
|
0.11
|
-0.31
|
-0.09
|
-0.05
|
-0.53
| 1 | | | | | | | |
|
P02
|
0.05
|
0.24
|
0.37
|
0.36
|
0.41
|
0.19
|
0.37
|
-0.42
|
-0.32
|
0.15
|
-0.42
| 0.75 | 1 | | | | | | |
|
P03
|
0.03
|
0.23
|
0.38
|
0.36
|
0.43
|
0.19
|
0.39
|
-0.46
|
-0.34
|
0.16
|
-0.46
| 0.76 | 0.99 | 1 | | | | | |
|
P04
|
-0.17
|
0.07
|
0.23
|
0.23
|
0.41
|
0.14
|
0.30
|
-0.56
|
-0.35
|
0.23
|
-0.61
| 0.78 | 0.82 | 0.86 | 1 | | | | |
|
P05
|
-0.32
|
-0.19
|
-0.13
|
-0.09
|
0.12
|
0.00
|
0.00
|
-0.34
|
-0.14
|
0.18
|
-0.44
|
0.32
|
0.15
|
0.19
| 0.77 | 1 | | | |
|
P06
|
-0.32
|
-0.19
|
-0.13
|
-0.09
|
0.13
|
0.00
|
0.00
|
-0.36
|
-0.15
|
0.19
|
-0.46
|
0.34
|
0.17
|
0.21
| 0.77 | 0.99 | 1 | | |
|
P07
|
-0.20
|
-0.1
|
-0.11
|
0.06
|
0.29
|
0.23
|
0.18
|
-0.45
|
-0.31
|
0.34
|
-0.37
|
0.14
|
0.31
|
0.33
| 0.75 | 0.91 | 0.85 | 1 | |
|
P08
|
0.55
|
0.56
|
0.57
|
0.56
|
0.34
|
0.29
|
0.47
|
0.00
|
-0.21
|
0.13
|
0.28
|
0.12
|
0.38
|
0.36
|
-0.41
|
-0.82
| -0.82 | -0.75 | 1 |
난대 목본식물종 별 분포지 및 한국 난대식생대의 공간 격자에서 선정된 기후 변수인 T07, T11 및 P04에 의한 MESS 분석에서 유사도 지수는 -604 - 69의 범위이었다. 이 유사도 지수에 영향을 미치는 주요 기후변수를 선별하기 위하여 단계적 변수선택 회귀분석을 실시한 결과 최종적으로 T07과 P04가 선별되었다 (Table 2). 선정된 변수의 설명력은 T07이 88.3%와 P04이 2.7%이었다 (Table 2). 따라서 난대 목본식물종 분포에는 최한 사분기 평균기온이 가장 중요한 기후요인임을 알 수 있었다. 분포의 유사도 지수 설명하는 회귀식은 y = 56.759 – 12.8116·T07 – 0.0073·P04로 추정되었다. 이 회귀식에 각 동아시아 난대 목본식물종 분포지의 평균 T07 및 P04를 입력하여 산출된 MESS 유사도 지수에서 0 이상이 4종이었고 나머지는 모두 음수로 나타났다 (Table 3 및 Appendix 1). 한국 난대식생대에 분포할 가능성이 가장 높은 종은 Lyonia villosa, Zanthoxylum bungeanum, Rhamnus globosa, Meliosma cuneifoliad이었고, 유사도 지수가 음수로서 비교적 높은 종은 Symplocos myrtacea, Celastrus hypoleucus, Wisteria sinensis, Rubus ichangensis, Smilax glaucochina, Elaeagnus henryi이었다.
Table 2.
Results of a stepwise regression (R2 = 0.89, p<0.001) estimating the multivariate environmental similarity surfaces (MESS) scores of the distribution of warm-temperate woody plants of East Asia in the warm-temperate vegetation zone of Korea according to major climate variables (n=25,016)
|
Climate variable
|
Coefficient
|
SE
| t | p |
Partial R2 |
|
Intercept
|
56.7579
|
0.5139
|
110.450
|
<0.001
|
-
|
|
Mean temperature of coldest quarter (T07)
|
-12.8116
|
0.0294
|
-435.360
|
<0.001
|
0.8834
|
|
Annual precipitation (P04)
|
-0.0073
|
0.0003
|
-26.620
|
<0.001
|
0.0275
|
Table 3.
List of warm-temperate woody plants of East Asia by the ranges of the scores from the multivariate environmental similarity surfaces (MESS) of mean temperature of the coldest quarter and annual precipitation between their distribution sites and the warm temperate vegetation zone of Korea. The score indicates the possibility of potential distribution of warm-temperate woody plants of East Asia in Korea
|
Score range
|
Species
|
No. of sp.
|
|
10 - 20
| Lyonia villosa, Zanthoxylum bungeanum, |
2
|
|
0 - 10
| Rhamnus globosa, Meliosma cuneifolia |
2
|
|
-10 - 0
| Symplocos myrtacea |
1
|
|
-20 - -10
| Celastrus hypoleucus, Wisteria sinensis |
2
|
|
-30 - -20
| Rubus ichangensis, Smilax glaucochina, Elaeagnus henryi |
3
|
|
-40 - -30
| Ulmus changii, Smilax menispermoidea, Rhamnus leptophylla, Rubus irenaeus, Celastrus gemmatus,
Sageretia henryi, Litsea coreana var. sinensis, Rubus malifolius, Symplocos dryophila, Acer campbellii,
Smilax polycolea, Eurya alata, Gaultheria griffithiana, Laurocerasus spinulosa |
14
|
|
-50 - -40
| Ilex corallina, Macropanax rosthornii, Lyonia ovalifolia, Osmanthus cooperi,
Phyllostachys heteroclada, Osteomeles schwerinae, Pleioblastus amarus, Phoebe
hunanensis, Styrax calvescens, Euonymus dielsianus, Castanopsis sclerophylla, Vaccinium
mandarinorum, Viburnum brachybotryum, Acer coriaceifolium, Parthenocissus laetevirens,
Camellia pitardii, Neolitsea aurata var. chekiangensis, Craibiodendron yunnanense,
Trachelospermum axillare, Clerodendrum mandarinorum |
20
|
|
-290 - -50
|
Not listed (Shown in Appendix 1)
|
275
|
|
Total
|
319
|
이상의 결과에서 본 연구에서, 중국과 일본에 분포하는 난대 식물의 한국 난대식생대에 잠재적으로 분포할 가능성을 평가하기 위하여, 생물기후 환경변수를 상관 분석을 실시하여 다중공선성을 배제하여 주요 환경변수로서 최한사분기 평균기온 (T07), 평균온도일교차 (T11) 및 연강수량 (P04)를 선별할 수 있었다. 또한 MESS 분석법을 산출한 한국 난대식생대와 각종별 분포지의 기후변수 유사도 지수에 영향을 미치는 기후변수 중에서 선택적 변수선택 회귀분석으로 최한사분기 평균기온 (T07)과 연강수량 (P04)을 선정하였다. 이 두 변수로 구성된 유사도 지수를 구할 수 있는 다중회귀식을 도출함으로써 동남아 난대 목본식물의 한국의 잠재분포 가능성을 평가할 수 있는 새로운 방법을 이 연구에서 제안할 수 있었다. 이 평가방법을 이용한다면 기후변화에 따라서 한국에 이입하여 정착할 수 있는 난대식물종을 예측하여 외래종 및 생태계 관리 대책을 수립할 수 있고, 적극적으로는 변화된 환경에서 생육이 적합한 식재종을 선발하는데 활용할 수 있을 것으로 기대된다.
Appendix
Appendix 1
Major climate variables and score indicating the potential distribution possibility estimated by the climate in the warm-temperate vegetation zone of Korea for warm-temperate plants distributed in East Asia not in Korea (mean±SD)
|
Species
|
n
|
Mean temperature of coldest
quarter (°C)
|
Annual precipitation (mm)
|
Score
|
| Lyonia villosa |
71
|
2.7±5.2
|
939±291
|
15.3
|
| Zanthoxylum bungeanum |
148
|
2.8±4.5
|
914±336
|
14.2
|
| Rhamnus globose |
56
|
3.5±4.2
|
1117±358
|
3.8
|
| Meliosma cuneifolia |
95
|
3.6±3.6
|
1012±275
|
3.3
|
| Symplocos myrtacea |
162
|
3.5±2.3
|
2474±507
|
-6.1
|
| Celastrus hypoleucus |
34
|
4.7±4.6
|
1184±373
|
-12.1
|
| Wisteria sinensis |
5234
|
5.3±2.7
|
866±256
|
-17.5
|
| Rubus ichangensis |
65
|
5.6±2.4
|
1247±213
|
-24.1
|
| Smilax glaucochina |
90
|
5.9±2.5
|
1348±294
|
-28.7
|
| Elaeagnus henryi |
60
|
6.0±2.6
|
1305±254
|
-29.6
|
| Ulmus changii |
12
|
6.0±2.3
|
1384±364
|
-30.2
|
| Smilax menispermoidea |
145
|
5.8±4.6
|
1770±989
|
-30.5
|
| Rhamnus leptophylla |
161
|
6.1±3.1
|
1249±281
|
-30.5
|
| Rubus irenaeus |
59
|
6.1±1.9
|
1378±200
|
-31.4
|
| Celastrus gemmatus |
110
|
6.3±2.9
|
1296±332
|
-33.4
|
| Sageretia henryi |
46
|
6.4±2.5
|
1281±211
|
-34.6
|
| Litsea coreana var. sinensis |
29
|
6.4±1.5
|
1470±189
|
-36.0
|
| Rubus malifolius |
54
|
6.5±3.5
|
1309±265
|
-36.1
|
| Symplocos dryophila |
50
|
6.6±5.6
|
1142±287
|
-36.1
|
| Acer campbellii |
37
|
6.7±5.0
|
1221±309
|
-38.0
|
| Smilax polycolea |
48
|
6.8±3.1
|
1235±272
|
-39.4
|
| Eurya alata |
73
|
6.7±2.1
|
1435±205
|
-39.5
|
| Gaultheria griffithiana |
54
|
6.9±4.9
|
1113±309
|
-39.8
|
| Laurocerasus spinulosa |
158
|
6.5±2.5
|
1818±424
|
-39.8
|
| Ilex corallina |
89
|
6.9±2.8
|
1184±253
|
-40.3
|
| Macropanax rosthornii |
50
|
6.8±2.6
|
1407±288
|
-40.6
|
| Lyonia ovalifolia |
613
|
6.5±4.2
|
2011±896
|
-41.2
|
| Osmanthus cooperi |
16
|
6.8±1.7
|
1494±231
|
-41.3
|
| Phyllostachys heteroclada |
15
|
6.9±3.0
|
1500±200
|
-42.6
|
| Osteomeles schwerinae |
63
|
7.3±4.6
|
836±208
|
-42.9
|
| Pleioblastus amarus |
25
|
7.1±2.4
|
1336±204
|
-44.0
|
| Phoebe hunanensis |
36
|
7.1±2.5
|
1480±131
|
-45.0
|
| Styrax calvescens |
21
|
7.1±2.1
|
1478±207
|
-45.0
|
| Euonymus dielsianus |
94
|
7.2±2.8
|
1331±222
|
-45.2
|
| Castanopsis sclerophylla |
145
|
7.2±2.4
|
1498±224
|
-46.4
|
| Vaccinium mandarinorum |
96
|
7.3±3.1
|
1386±290
|
-46.9
|
| Viburnum brachybotryum |
93
|
7.4±3.0
|
1326±227
|
-47.7
|
| Acer coriaceifolium |
12
|
7.4±1.7
|
1329±184
|
-47.7
|
| Parthenocissus laetevirens |
32
|
7.4±3.2
|
1405±330
|
-48.3
|
| Camellia pitardii |
68
|
7.5±2.4
|
1239±263
|
-48.4
|
| Neolitsea aurata var. chekiangensis |
24
|
7.4±2.6
|
1453±322
|
-48.6
|
| Craibiodendron yunnanense |
28
|
7.6±4.7
|
1178±345
|
-49.2
|
| Trachelospermum axillare |
126
|
7.5±4.0
|
1386±297
|
-49.4
|
| Clerodendrum mandarinorum |
88
|
7.5±3.1
|
1427±202
|
-49.7
|
| Phoebe sheareri |
139
|
7.5±2.2
|
1532±189
|
-50.5
|
| Euonymus vagans |
59
|
7.7±5.0
|
1215±352
|
-50.8
|
| Rosa cymosa |
232
|
7.6±3.1
|
1397±326
|
-50.8
|
| Rubus paniculatus |
19
|
7.7±6.3
|
1280±432
|
-51.2
|
| Uncaria rhynchophylla |
125
|
7.4±3.4
|
1883±471
|
-51.8
|
| Prunus spinulosa |
134
|
7.3±4.2
|
2067±622
|
-51.8
|
| Rhododendron leptothrium |
30
|
7.9±4.5
|
1098±287
|
-52.5
|
| Zanthoxylum echinocarpum |
31
|
7.8±2.1
|
1361±140
|
-53.1
|
| Acer cordatum |
77
|
7.8±3.0
|
1545±271
|
-54.4
|
| Photinia parvifolia |
165
|
7.8±2.6
|
1564±332
|
-54.6
|
| Illicium simonsii |
45
|
8.1±4.4
|
1051±326
|
-54.7
|
| Rosa longicuspis |
96
|
8.1±4.0
|
1077±281
|
-54.9
|
| Machilus yunnanensis |
21
|
8.3±4.3
|
1026±276
|
-57.1
|
| Symplocos ramosissima |
88
|
8.2±4.0
|
1344±313
|
-58.1
|
| Ilex hylonoma |
15
|
8.2±2.4
|
1449±164
|
-58.9
|
| Rhododendron mariesii |
208
|
8.1±3.6
|
1735±713
|
-59.7
|
| Litsea wilsonii |
36
|
8.4±5.4
|
1259±286
|
-60.0
|
| Hypericum uralum |
19
|
8.3±7.2
|
1488±705
|
-60.4
|
| Piper wallichii |
46
|
8.4±5.6
|
1408±460
|
-61.1
|
| Lyonia ovalifolia var. lanceolata |
104
|
8.5±4.5
|
1223±434
|
-61.1
|
| Alnus nepalensis |
159
|
8.5±4.8
|
1283±431
|
-61.5
|
| Ficus henryi |
79
|
8.5±4.2
|
1342±283
|
-61.9
|
| Lithocarpus glaber |
247
|
8.3±3.3
|
1755±377
|
-62.4
|
| Smilax ferox |
103
|
8.6±4.1
|
1250±286
|
-62.5
|
| Ormosia henryi |
37
|
8.5±2.6
|
1543±181
|
-63.4
|
| Cinnamomum appelianum |
42
|
8.6±2.5
|
1488±229
|
-64.3
|
| Rhododendron ovatum |
200
|
8.4±2.9
|
1855±565
|
-64.4
|
| Millettia dielsiana |
126
|
8.7±3.9
|
1411±292
|
-65.0
|
| Mahonia duclouxiana |
16
|
8.9±3.5
|
1100±234
|
-65.3
|
| Schima argentea |
102
|
8.8±3.4
|
1332±315
|
-65.7
|
| Eurya jintungensis |
15
|
8.8±3.8
|
1347±274
|
-65.8
|
| Elaeocarpus glabripetalus |
78
|
8.7±3.1
|
1556±188
|
-66.1
|
| Pistacia chinensis |
1655
|
9.0±3.6
|
1035±517
|
-66.1
|
| Vaccinium duclouxii |
67
|
9.0±3.2
|
1113±266
|
-66.7
|
| Laurocerasus zippeliana |
107
|
8.6±3.2
|
1879±548
|
-67.1
|
| Tetrastigma serrulatum |
69
|
8.9±4.8
|
1382±799
|
-67.3
|
| Kadsura longipedunculata |
51
|
8.8±2.6
|
1553±170
|
-67.3
|
| Styrax odoratissimus |
31
|
8.8±3.4
|
1555±161
|
-67.3
|
| Actinidia eriantha |
102
|
8.8±2.2
|
1580±172
|
-67.5
|
| Castanopsis eyrei |
246
|
8.8±3.1
|
1655±356
|
-68.1
|
| Eurya rubiginosa var. attenuata |
68
|
8.9±2.7
|
1576±144
|
-68.8
|
| Stauntonia chinensis |
22
|
9.0±5.7
|
1452±385
|
-69.1
|
| Symplocos lancifolia |
357
|
8.7±3.8
|
2047±675
|
-69.6
|
| Ficus sarmentosa var. henryi |
259
|
8.9±4.1
|
1766±709
|
-70.1
|
| Leucosceptrum canum |
57
|
9.1±6.0
|
1438±580
|
-70.3
|
| Symplocos stellaris |
356
|
8.7±3.4
|
2392±822
|
-72.1
|
| Rubus columellaris |
66
|
9.2±2.9
|
1534±172
|
-72.3
|
| Rhododendron microphyton |
77
|
9.4±4.1
|
1205±310
|
-72.5
|
| Isodon amethystoides |
15
|
9.2±4.5
|
2027±915
|
-75.9
|
| Sloanea sinensis |
141
|
9.5±3.6
|
1557±265
|
-76.3
|
| Jasminum lanceolaria |
248
|
9.5±4.6
|
1585±458
|
-76.5
|
| Ampelopsis grossedentata |
93
|
9.6±2.9
|
1541±198
|
-77.5
|
| Symplocos heishanensis |
142
|
9.1±2.9
|
2627±637
|
-79.0
|
| Lindera aggregata |
236
|
9.6±3.9
|
1804±476
|
-79.4
|
| Ficus pandurata |
85
|
9.8±3.5
|
1561±219
|
-80.2
|
| Meliosma rigida |
195
|
9.3±4.0
|
2465±539
|
-80.4
|
| Tetrastigma hemsleyanum |
126
|
9.8±3.5
|
1830±601
|
-82.1
|
| Premna microphylla |
268
|
9.7±4.1
|
2158±751
|
-83.3
|
| Aucuba chlorascens |
14
|
10.2±4.8
|
1372±349
|
-83.9
|
| Symplocos anomala |
221
|
9.9±5.8
|
1890±743
|
-83.9
|
| Alangium chinense |
358
|
10.2±6.3
|
1458±473
|
-84.6
|
| Embelia vestita |
44
|
10.2±5.9
|
1473±631
|
-84.7
|
| Choerospondias axillaris |
127
|
10.2±4.4
|
1474±281
|
-84.7
|
| Tarenna mollissima |
96
|
10.2±3.4
|
1617±213
|
-85.7
|
| Piper hancei |
73
|
10.3±4.2
|
1602±263
|
-86.9
|
| Camellia cordifolia |
52
|
10.3±2.2
|
1606±148
|
-86.9
|
| Castanopsis fargesii |
258
|
10.2±3.4
|
1799±449
|
-87.0
|
| Symplocos sumuntia |
182
|
10.2±5.6
|
1787±747
|
-87.0
|
| Smilax glabra |
237
|
10.3±4.2
|
1713±533
|
-87.7
|
| Callicarpa rubella |
259
|
10.6±4.3
|
1578±473
|
-90.6
|
| Castanopsis fordii |
71
|
10.6±2.5
|
1619±147
|
-90.9
|
| Lithocarpus hancei |
226
|
10.5±3.9
|
1861±720
|
-91.3
|
| Syzygium austrosinense |
15
|
10.7±3.7
|
1544±194
|
-91.6
|
| Dalbergia hancei |
119
|
10.7±3.9
|
1581±270
|
-91.9
|
| Ardisia hanceana |
42
|
10.8±4.1
|
1656±207
|
-93.7
|
| Coptosapelta diffusa |
173
|
10.7±3.5
|
1907±575
|
-94.2
|
| Tarenna depauperata |
26
|
11.0±2.4
|
1516±193
|
-95.2
|
| Eurya loquaiana |
873
|
10.4±3.4
|
2644±768
|
-95.8
|
| Dodonaea viscosa |
9246
|
11.5±3.7
|
828±511
|
-96.6
|
| Machilus pauhoi |
37
|
11.0±4.1
|
1748±324
|
-96.9
|
| Litsea cubeba |
910
|
10.9±4.7
|
2050±670
|
-97.8
|
| Castanopsis chinensis |
41
|
11.2±3.2
|
1627±405
|
-98.6
|
| Rhododendron mariae |
50
|
11.3±2.3
|
1583±194
|
-99.6
|
| Olea europaea subsp. cuspidata |
3028
|
11.8±1.4
|
926±217
|
-101.2
|
| Castanopsis echinocarpa |
12
|
11.5±4.3
|
1490±312
|
-101.4
|
| Sabia swinhoei |
125
|
11.1±3.6
|
2216±747
|
-101.6
|
| Rubus swinhoei |
230
|
11.0±4.0
|
2438±765
|
-102.0
|
| Alchornea trewioides |
108
|
11.5±3.7
|
1639±310
|
-102.5
|
| Psychotria calocarpa |
26
|
11.6±6.6
|
1484±500
|
-102.7
|
| Actinodaphne obovata |
11
|
11.5±3.5
|
1742±784
|
-103.3
|
| Neolitsea chuii |
59
|
11.6±2.3
|
1664±152
|
-104.0
|
| Meliosma arnottiana |
22
|
11.8±7.0
|
1369±366
|
-104.4
|
| Antidesma japonicum |
240
|
11.6±4.6
|
1717±480
|
-104.4
|
| Symplocos glauca |
384
|
11.1±4.0
|
2608±649
|
-104.5
|
| Prunus zippeliana |
109
|
11.4±4.3
|
2269±440
|
-105.8
|
| Alyxia sinensis |
66
|
11.8±3.1
|
1627±224
|
-106.3
|
| Boehmeria clidemioides |
57
|
11.9±7.1
|
1468±513
|
-106.4
|
| Lindera communis |
663
|
11.7±4.1
|
2142±852
|
-108.8
|
| Litsea acuminata |
923
|
11.4±3.0
|
2751±540
|
-109.4
|
| Ilex pubescens |
301
|
11.8±3.2
|
2109±650
|
-109.8
|
| Helicia cochinchinensis |
362
|
11.7±3.7
|
2294±664
|
-109.9
|
| Symplocos viridissima |
18
|
12.3±5.6
|
1543±244
|
-112.1
|
| Smilax lanceifolia |
343
|
11.8±4.0
|
2456±706
|
-112.3
|
| Ficus cyrtophylla |
67
|
12.4±4.7
|
1417±291
|
-112.4
|
| Elaeocarpus varunua |
17
|
12.4±5.8
|
1531±327
|
-113.3
|
| Celastrus hindsii |
142
|
12.2±4.3
|
1932±755
|
-113.6
|
| Castanopsis carlesii |
230
|
11.9±4.0
|
2451±744
|
-113.6
|
| Phoebe puwenensis |
13
|
12.6±6.2
|
1403±362
|
-114.9
|
| Ficus variolosa |
146
|
12.5±3.4
|
1690±276
|
-115.7
|
| Ampelopsis cantoniensis |
244
|
12.3±3.8
|
2101±649
|
-116.1
|
| Gnetum parvifolium |
53
|
12.6±3.8
|
1667±383
|
-116.8
|
| Leea compactiflora |
13
|
12.8±6.8
|
1450±388
|
-117.8
|
| Smilax hypoglauca |
32
|
12.7±2.9
|
1643±268
|
-117.9
|
| Castanopsis hystrix |
171
|
12.6±4.0
|
1891±546
|
-118.5
|
| Lindera chunii |
22
|
12.7±4.0
|
1752±329
|
-118.7
|
| Ilex viridis |
57
|
12.7±5.2
|
1800±435
|
-119.1
|
| Syzygium buxifolium |
462
|
12.4±3.6
|
2414±738
|
-119.7
|
| Myriopteron extensum |
14
|
13.0±4.0
|
1377±215
|
-119.8
|
| Diospyros morrisiana |
145
|
12.5±2.6
|
2315±654
|
-120.3
|
| Lindera megaphylla |
289
|
12.5±3.8
|
2462±777
|
-121.3
|
| Cleidion brevipetiolatum |
46
|
13.1±4.7
|
1419±290
|
-121.4
|
| Anodendron affine |
154
|
12.6±4.9
|
2338±567
|
-121.7
|
| Castanopsis fissa |
161
|
13.0±3.5
|
1746±292
|
-122.5
|
| Calamus thysanolepis |
15
|
13.1±4.0
|
1607±146
|
-122.8
|
| Camellia furfuracea |
41
|
13.0±3.7
|
1888±462
|
-123.6
|
| Fissistigma oldhamii |
243
|
12.8±3.0
|
2295±596
|
-124.0
|
| Aidia cochinchinensis |
213
|
12.9±4.6
|
2210±720
|
-124.6
|
| Toona ciliata |
1873
|
13.5±3.4
|
1341±394
|
-126.0
|
| Dioscorea cirrhosa |
154
|
13.0±4.6
|
2252±746
|
-126.2
|
| Ficus semicordata |
66
|
13.5±5.5
|
1461±412
|
-126.9
|
| Myrsine seguinii |
606
|
12.7±3.5
|
2883±583
|
-127.0
|
| Erythrophleum fordii |
12
|
13.6±5.9
|
1613±420
|
-129.2
|
| Eurya trichocarpa |
44
|
13.5±6.6
|
1864±560
|
-129.8
|
| Litsea verticillata |
56
|
13.7±4.0
|
1608±293
|
-130.5
|
| Eleutherococcus trifoliatus |
163
|
13.4±3.5
|
2265±537
|
-131.4
|
| Anneslea fragrans |
66
|
13.9±5.0
|
1439±321
|
-131.8
|
| Sarcosperma arboreum |
31
|
13.9±5.0
|
1601±1111
|
-133.0
|
| Randia cochinchinensis |
389
|
13.3±3.2
|
2838±573
|
-134.3
|
| Mussaenda erosa |
61
|
14.0±4.2
|
1627±304
|
-134.5
|
| Castanopsis fleuryi |
12
|
14.2±2.2
|
1364±228
|
-135.1
|
| Ardisia lindleyana |
43
|
13.9±2.3
|
1934±192
|
-135.4
|
| Mussaenda pubescens |
202
|
14.0±3.3
|
1903±355
|
-136.5
|
| Lithocarpus truncatus |
61
|
14.2±3.7
|
1653±333
|
-137.2
|
| Tetrastigma planicaule |
52
|
14.3±3.8
|
1615±365
|
-138.2
|
| Gnetum montanum |
62
|
14.5±6.3
|
1613±599
|
-140.8
|
| Symplocos racemosa |
70
|
14.5±6.9
|
1625±781
|
-140.9
|
| Machilus chinensis |
22
|
14.6±5.7
|
1575±206
|
-141.8
|
| Lasianthus fordii |
628
|
13.9±2.6
|
2841±549
|
-142.0
|
| Blastus cochinchinensis |
636
|
14.0±2.3
|
2768±516
|
-142.8
|
| Albizia lucidior |
14
|
14.7±3.6
|
1552±404
|
-142.9
|
| Eurya nitida |
208
|
14.0±5.0
|
2788±586
|
-142.9
|
| Engelhardia roxburghiana |
659
|
14.0±2.6
|
2867±577
|
-143.5
|
| Cryptocarya chinensis |
318
|
14.2±2.2
|
2653±606
|
-144.5
|
| Mallotus philippensis |
3158
|
14.9±4.4
|
1574±580
|
-145.6
|
| Tarenna attenuata |
35
|
14.9±4.5
|
1581±290
|
-145.7
|
| Lithocarpus dealbatus |
125
|
15.0±6.1
|
1466±618
|
-146.1
|
| Magnolia henryi |
17
|
15.0±3.8
|
1492±246
|
-146.3
|
| Litsea monopetala |
109
|
14.9±6.2
|
1675±515
|
-146.4
|
| Pericampylus glaucus |
249
|
14.7±5.6
|
2110±761
|
-147.0
|
| Pinus kesiya |
54
|
15.0±4.0
|
1619±685
|
-147.2
|
| Ilex asprella |
566
|
14.4±2.1
|
2703±575
|
-147.4
|
| Byttneria aspera |
29
|
15.0±2.6
|
1759±282
|
-148.2
|
| Ardisia quinquegona |
593
|
14.5±2.3
|
2640±611
|
-148.3
|
| Bauhinia championii |
231
|
14.7±2.6
|
2338±542
|
-148.6
|
| Dasymaschalon rostratum |
11
|
15.3±5.0
|
1528±139
|
-150.4
|
| Castanopsis argyrophylla |
41
|
15.4±4.5
|
1563±430
|
-151.9
|
| Cinnamomum bejolghota |
78
|
15.3±5.7
|
1773±617
|
-152.2
|
| Radermachera sinica |
176
|
15.0±3.8
|
2343±722
|
-152.5
|
| Lasianthus curtisii |
95
|
14.8±2.9
|
2703±480
|
-152.6
|
| Fissistigma glaucescens |
33
|
15.3±4.1
|
1916±573
|
-153.2
|
| Cryptolepis buchananii |
56
|
15.5±5.0
|
1697±840
|
-154.2
|
| Sterculia lanceolata |
224
|
15.4±3.0
|
1904±291
|
-154.4
|
| Schefflera heptaphylla |
692
|
15.1±1.9
|
2470±495
|
-154.7
|
| Garcinia oblongifolia |
52
|
15.5±2.7
|
1834±250
|
-155.2
|
| Psychotria serpens |
562
|
15.1±2.6
|
2545±560
|
-155.3
|
| Croton tiglium |
135
|
15.6±6.7
|
1730±592
|
-155.7
|
| Schima noronhae |
13
|
15.3±4.9
|
2266±615
|
-155.8
|
| Smilax ocreata |
94
|
15.5±4.3
|
2004±670
|
-156.4
|
| Garcinia cowa |
18
|
15.8±2.7
|
1504±266
|
-156.6
|
| Melastoma candidum |
631
|
15.1±2.1
|
2735±538
|
-156.6
|
| Symplocos microcalyx |
24
|
15.4±1.1
|
2313±195
|
-157.4
|
| Trema angustifolia |
45
|
15.7±5.9
|
1845±686
|
-157.8
|
| Canarium album |
38
|
15.8±4.3
|
1676±331
|
-157.9
|
| Bredia hirsuta |
21
|
15.4±4.4
|
2436±390
|
-158.3
|
| Barleria cristata |
284
|
16.0±6.2
|
1460±670
|
-158.9
|
| Mussaenda parviflora |
169
|
15.4±3.1
|
2544±490
|
-159.1
|
| Ardisia sieboldii |
1128
|
15.3±2.2
|
2892±507
|
-160.4
|
| Ficus auriculata |
176
|
16.1±6.6
|
1609±738
|
-161.2
|
| Dendrolobium triangulare |
107
|
16.2±5.8
|
1692±587
|
-163.1
|
| Caryota maxima |
16
|
16.1±3.8
|
2003±598
|
-164.1
|
| Callicarpa nudiflora |
31
|
16.3±5.2
|
1662±410
|
-164.2
|
| Alphonsea monogyna |
19
|
16.5±4.5
|
1467±262
|
-165.3
|
| Tarennoidea wallichii |
59
|
16.4±6.3
|
1670±563
|
-165.5
|
| Ilex godajam |
14
|
16.4±5.3
|
1803±598
|
-166.5
|
| Melastoma sanguineum |
134
|
16.4±3.9
|
1919±366
|
-167.3
|
| Pittosporopsis kerrii |
16
|
16.5±2.8
|
1810±415
|
-167.8
|
| Maesa indica |
69
|
16.5±6.8
|
1800±896
|
-167.8
|
| Diospyros eriantha |
721
|
16.0±2.3
|
2693±469
|
-167.9
|
| Terminalia myriocarpa |
24
|
16.6±4.6
|
1732±658
|
-168.6
|
| Tetracera sarmentosa |
20
|
16.5±4.2
|
2020±355
|
-169.4
|
| Rhodomyrtus tomentosa |
452
|
16.5±4.0
|
2025±598
|
-169.4
|
| Rubus alceifolius |
144
|
16.3±3.5
|
2393±922
|
-169.5
|
| Osmanthus marginatus |
257
|
16.1±3.9
|
2770±768
|
-169.7
|
| Castanopsis indica |
216
|
16.4±3.2
|
2273±816
|
-169.9
|
| Aporosa yunnanensis |
17
|
16.8±4.8
|
1610±428
|
-170.2
|
| Schima wallichii |
266
|
16.7±5.7
|
1986±806
|
-171.7
|
| Triadica cochinchinensis |
151
|
16.4±3.7
|
2572±652
|
-172.1
|
| Tabernaemontana corymbosa |
24
|
16.8±5.5
|
2033±578
|
-173.3
|
| Cryptocarya concinna |
205
|
16.5±2.9
|
2703±634
|
-174.3
|
| Ilex warburgii |
34
|
16.7±1.9
|
2342±526
|
-174.3
|
| Alpinia zerumbet |
1511
|
17.0±3.9
|
1984±727
|
-175.5
|
| Tarenna gracilipes |
210
|
16.7±3.0
|
2725±508
|
-177.1
|
| Stereospermum colais |
22
|
17.4±4.1
|
1509±446
|
-177.2
|
| Phyllanthus emblica |
298
|
17.4±6.0
|
1646±692
|
-178.2
|
| Psychotria henryi |
11
|
17.5±2.2
|
1564±176
|
-178.9
|
| Ficus microcarpa |
1644
|
17.5±4.6
|
1872±719
|
-181.1
|
| Lasianthus trichophlebus |
15
|
18.0±4.4
|
1576±326
|
-185.3
|
| Engelhardia spicata |
27
|
17.7±5.2
|
2295±718
|
-186.7
|
| Celastrus paniculatus |
98
|
18.1±5.0
|
1619±640
|
-186.9
|
| Carissa spinarum |
4126
|
18.6±4.8
|
816±361
|
-187.5
|
| Duabanga grandiflora |
65
|
18.1±4.6
|
1772±682
|
-188.1
|
| Lasianthus verticillatus |
79
|
17.8±3.4
|
2581±616
|
-190.1
|
| Psychotria manillensis |
30
|
18.0±1.5
|
2294±257
|
-190.6
|
| Antidesma pentandrum |
11
|
18.1±0.8
|
2142±81
|
-190.8
|
| Canthium horridum |
59
|
18.3±5.9
|
1891±587
|
-191.5
|
| Antidesma acidum |
51
|
18.4±5.9
|
1741±930
|
-191.7
|
| Woodfordia fruticosa |
104
|
18.6±5.8
|
1751±1016
|
-194.3
|
| Vitex quinata |
270
|
18.3±5.2
|
2479±540
|
-195.8
|
| Ficus ampelas |
322
|
18.3±4.1
|
2643±487
|
-197.0
|
| Acronychia pedunculata |
200
|
18.5±4.8
|
2303±735
|
-197.1
|
| Caesalpinia crista |
176
|
18.7±5.8
|
2118±780
|
-198.3
|
| Cipadessa baccifera |
35
|
19.1±4.0
|
1485±681
|
-198.8
|
| Dendrotrophe varians |
72
|
18.8±5.8
|
2381±715
|
-201.5
|
| Diploclisia glaucescens |
47
|
18.9±4.8
|
2407±990
|
-202.9
|
| Cajanus scarabaeoides |
393
|
19.5±4.7
|
1644±720
|
-205.1
|
| Crotalaria linifolia |
121
|
19.5±6.0
|
1676±590
|
-205.3
|
| Dalbergia pinnata |
94
|
19.6±7.2
|
2263±908
|
-210.9
|
| Trema tomentosa |
941
|
20.3±4.9
|
1410±662
|
-213.6
|
| Cratoxylum cochinchinense |
139
|
20.1±5.8
|
2160±695
|
-216.5
|
| Excoecaria agallocha |
620
|
20.6±4.5
|
1744±652
|
-219.9
|
| Cerbera manghas |
520
|
20.4±3.8
|
2177±799
|
-220.5
|
| Baccaurea ramiflora |
121
|
20.7±3.8
|
1731±632
|
-221.1
|
| Lasianthus cyanocarpus |
68
|
20.3±4.0
|
2563±543
|
-222.0
|
| Phoebe lanceolata |
50
|
20.6±4.3
|
2041±815
|
-222.1
|
| Melicope triphylla |
90
|
20.2±3.0
|
2760±642
|
-222.2
|
| Barringtonia racemosa |
363
|
20.5±4.5
|
2249±948
|
-222.3
|
| Entada phaseoloides |
146
|
20.5±4.7
|
2455±798
|
-223.8
|
| Smilax zeylanica |
26
|
20.9±5.2
|
2212±653
|
-227.1
|
| Melastoma malabathricum |
939
|
21.0±5.7
|
2351±731
|
-229.4
|
| Decaspermum fruticosum |
81
|
21.1±5.1
|
2373±808
|
-230.9
|
| Lannea coromandelica |
62
|
21.4±4.6
|
2010±1058
|
-232.1
|
| Ficus virens |
1166
|
21.9±3.9
|
1392±472
|
-234.0
|
| Bridelia stipularis |
139
|
21.6±6.0
|
2115±815
|
-235.4
|
| Bridelia tomentosa |
1563
|
21.9±3.7
|
1615±601
|
-235.6
|
| Ficus fulva |
72
|
21.6±5.5
|
2571±725
|
-238.7
|
| Antidesma montanum |
264
|
21.8±5.5
|
2264±723
|
-239.1
|
| Tephrosia purpurea |
1164
|
22.7±3.2
|
729±630
|
-239.4
|
| Heritiera littoralis |
279
|
21.9±3.3
|
2188±751
|
-239.8
|
| Rourea minor |
194
|
22.0±3.8
|
2064±731
|
-240.2
|
| Gironniera subaequalis |
145
|
21.9±4.7
|
2339±719
|
-240.9
|
| Ficus subulata |
261
|
22.0±4.4
|
2618±684
|
-244.2
|
| Hernandia nymphaeifolia |
124
|
22.3±3.1
|
2249±800
|
-245.3
|
| Syzygium acuminatissimum |
31
|
22.6±3.9
|
2374±267
|
-250.1
|
| Leea indica |
186
|
22.7±4.5
|
2431±794
|
-251.8
|
| Knema furfuracea |
20
|
22.8±3.2
|
2316±443
|
-252.2
|
| Knema globularia |
53
|
23.0±4.1
|
1999±607
|
-252.5
|
| Microdesmis caseariifolia |
71
|
23.3±4.1
|
2023±544
|
-256.5
|
| Poikilospermum suaveolens |
53
|
23.1±4.1
|
2402±643
|
-256.7
|
| Diospyros maritima |
579
|
23.8±2.2
|
1569±538
|
-259.6
|
| Lepisanthes senegalensis |
170
|
24.6±3.5
|
1399±781
|
-268.6
|
| Pometia pinnata |
204
|
24.1±2.5
|
2654±780
|
-271.4
|
| Antiaris toxicaria |
3002
|
25.1±1.2
|
1165±316
|
-273.3
|
| Orophea creaghii |
30
|
24.5±3.9
|
2365±552
|
-274.4
|
| Barringtonia macrostachya |
34
|
24.6±3.2
|
2475±619
|
-276.5
|
| Walsura pinnata |
21
|
25.6±1.5
|
2577±491
|
-290.0
|
Acknowledgements
본 연구는 국립산림과학원 난대·아열대연구소 ‘도서 해안지역 탄소흡수원 확충을 위한 맹그로브 적응성 검증 및 조성기반 구축 연구’ (과제번호 FE0100-2022-04-2022)에 의해 수행되었습니다.
References
Aho, K. 2022. asbio: A collection of statistical tools for biologists, R package version 1.8-3. https://CRAN.R-project.org/package=asbio. Accessed 4 December 2022.
Barbosa, A.M., Real, R., Munoz, A.R., and Brown, J.A. 2013. New measures for assessing model equilibrium and prediction mismatch in species distribution models. Diversity and Distributions 19: 1333-1338.
10.1111/ddi.12100Cho, K.H. and Lee, S.H. 2015. Prediction of changes in the potential distribution of a waterfront alien plant,
Paspalum distichum var.
indutum, under climate change in the Korean Peninsula. Ecology and Resilient Infrastructure 2: 206-215.
10.17820/eri.2015.2.3.206Elith, J. and Leathwick, J.R. 2009. Species distribution models: ecological explanation and prediction across space and time. Annual Review of Ecology, Evolution and Systematics 40: 677-697.
10.1146/annurev.ecolsys.110308.120159Elith, J., Kearney, M. and Phillips, S. 2010. The art of modelling range‐shifting species. Methods in Ecology and Evolution 1: 330-342.
10.1111/j.2041-210X.2010.00036.xGBIF. 2022. Free and open access to biodiversity data. Global Biodiversity Information Facility Secretariat. https://www.gbif.org. Accessed 24 April 2021.
IPCC. 2021. Climate change 2021: The physical science basis. Intergovernmental Panel on Climate Change, Geneva, Switzerland.
Kira, T. 1945. A new classification of climate in eastern Asia as the basis for agricultural geography. Horticultural Institute, Kyoto University, Kyoto, Japan. (In Japanese)
Kira, T. 1991. Forest ecosystems of east and southeast Asia in a global perspective. Ecological Research 6(2): 185-200.
10.1007/BF02347161KMA. 2021. Korea’s 109 climate change analysis report. Korea Meteorological Administration, Seoul, South Korea. (in Korean)
Mesgaran, M.B., Cousens, R.D. and Webber, B.L. 2014. Here be dragons: a tool for quantifying novelty due to covariate range and correlation change when projecting species distribution models. Diversity and Distributions 20: 1147-1159.
10.1111/ddi.12209Negrini, M., Fidelis, E.G., Picanço, M.C. and Ramos, R.S. 2020. Mapping of the
Steneotarsonemus spinki invasion risk in suitable areas for rice (
Oryza sativa) cultivation using MaxEnt. Experimental and Applied Acarology 80: 445-461.
10.1007/s10493-020-00474-632072353NIFS. 2012. Impact assessment and monitoring for terrestrial ecosystem to climate change on Juju Island. National Institute of Forest Science, Seoul, South Korea. (In Korean)
NIFS. 2021. Analysis of potential distribution area of subtropical forest life resources in response to climate change. National Institute of Forest Science, Seoul, South Korea. (in Korean)
NIFS. 2022. Analysis and prediction of potential growth sites of Mangroves in Korea. National Institute of Forest Science, Seoul, South Korea. (in Korean)
Oksanen, J., Simpson, G., Blanchet, F., Kindt, R., Legendre, P., Minchin, P., ... and Weedon, J. 2022. Vegan: Community ecology package, R package, version 2.6-2. https://CRAN.R-project.org/package=vegan.
Owens, H.L., Campbell, L.P., Dornak, L.L., Saupe, E.E., Barve, N., Soberon, J., Ingenloff, K., Lira-Noriega, A., Hensz, C.M., Myers, C.E., and Peterson, A.T. 2013. Constraints on interpretation of ecological niche models by limited environmental ranges on calibration areas. Ecological Modelling 263: 10-18.
10.1016/j.ecolmodel.2013.04.011Padalia, H., Srivastava, V., and Kushwaha, S.P.S. 2014. Modeling potential invasion range of alien invasive species,
Hyptis suaveolens (L.) Poit. in India: Comparison of MaxEnt and GARP. Ecological Informatics 22: 36-43.
10.1016/j.ecoinf.2014.04.002QGIS.org. 2022. QGIS Geographic Information System. QGIS Association. http://www.qgis.org. Accessed 20 June 2022.
R Core Team. 2022. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Santini, L., Benítez‐López, A., Maiorano, L., Čengić, M. and Huijbregts, M.A. 2021. Assessing the reliability of species distribution projections in climate change research. Diversity and Distributions 27: 1035-1050.
10.1111/ddi.13252Wang, H., Harrison, S.P., Prentice, I.C., Yang, Y., Bai, F., Togashi, H.F., Wang, M., Zhou, S., and Ni, J. 2018. The China plant trait database: Toward a comprehensive regional compilation of functional traits for land plants. Ecology 99: 500.
10.1002/ecy.209129155446WFO. 2022. An online flora of all known plants. World Flora Online. http://www.worldfloraonline.org. Accessed 4 April 2022.
WorldClim. 2022. Global climate data. WorldClim. https://www.worldclim.org. Accessed 1 November 2022.
WWF. 2015. Impact of climate change on species. World Wide Fund for Nature, Geneva, Switzerland.
Yim, Y.J. and Kira, T. 1975. Distribution of forest vegetation and climate in the Korean Peninsula. I. Distribution of some indices of thermal climate. Japanese Journal of Ecology 25(2): 77-88.


