

1. 서 론
2. 연구방법
2.1 조사지 개황
2.2 어류 채집
2.3 환경 조사
2.4 자료 분석
3. 결 과
3.1 환경 특성
3.2 환경과 조사지점의 관계
3.3 어류상
3.4 어류 군집 구조
3.5 어류 군집 구조와 환경과의 관계
4. 고 찰
5. 결 론
1. 서 론
하천 경관의 공간적 규모에서 3 차원을 따라서 종적, 횡적 및 수직적 연결성이 생성되고 변형된다 (Ward 1989). 하천 생태계에서 연속성이란 수문적 경관을 가로질러 물이 매개하는 에너지, 물질 및 생물의 이동을 의미한다 (Pringle 2003). 특히 하천의 횡적 연결성 (lateral connectivity)은 하천 하도와 이와 연결된 홍수터 (floodplain) 사이의 관계를 개념화한 것이다. 즉 하도-홍수터 공간은 역동적인 공간 모자이크로서 횡적 물 흐름이 다양한 경관 조각 (patch) 사이의 연결에 중요하다 (Thoms 2003).
하도와 홍수터에서 범람 파동 개념 (flood pulse concept)은 횡적 연결성의 생태적 중요성을 강조한다 (Sparks 1995). 이 개념에 따르면 하천 생물의 생활사는 하도와 홍수터에서 주기적인 수위 변동과 연관되어 있으며 (Kwak 1988, Miranda 2005), 특히 특정한 어류의 생활사는 본류와 홍수터의 연결성에 의하여 큰 영향을 받는다 (Kingsford 2000). 이들 어류는 하도의 수위가 높아짐에 따라서 범람된 홍수터로 이동하고 이곳에서 산란, 치어 성장 및 피난 은신하고 다시 수위가 낮아지면 본류로 돌아가서 생활사를 잇는다 (Rose and Baker 1983, Turner et al. 1994). 또한 하도와 연결성이 확보된 홍수터는 생산성과 생물다양성이 가장 높은 생태계 중의 하나이다. 이곳은 영양소, 퇴적물 및 유기물의 재분배와 순환, 수질 유지, 다양한 생물종에게 서식지와 먹이 공급 등의 중요한 생태계 기능을 제공하다 (Junk et al. 1989).
우리나라의 대부분 하천에서는 홍수터의 토지이용과 홍수 방재를 위한 하천정비에 의하여 인공제방을 축조하였다. 이러한 결과 제내지의 홍수터 대부분이 농경지, 주택지 등으로 이용되고 있으며, 잔존하는 홍수터와 구하도 (former floodplain)마저 하도로부터 격리되어 하천의 연결성이 단절되었다. 홍수터와 구하도가 본류 하도와 단절되면 하천 환경의 구조적 변화를 유발하여 홍수터뿐만 아니라 본류의 생물 서식처 다양성을 감소시키고 어류의 종 다양성까지 감소시킨다 (Schiemer 2000, Miyazono et al. 2010). 특히 홍수터가 하도로부터 단절되면 수질 악화 및 토사 퇴적이 발생하여 생물 서식지에 악영향을 미친다 (Miranda et al. 2001). 또한 오랫동안 격리된 홍수터와 구하도는 본류와 전혀 다른 환경으로 변하고 이에 따라 이곳의 어류 군집은 본류와는 크게 다른 특성을 보이게 된다 (Erskine et al. 2005, Andrews et al. 2014).
홍수터 또는 구하도가 본류 하도와 단절된 정비 하천에서는 하천 생태계의 건강성을 유지하고 개선하기 위하여 횡적 연결성을 확보하는 것이 매우 중요하다 (Tockner et al. 1999, Aarts et al. 2004). 하천의 횡적 연결성을 유지, 복원하기 위하여 하도-홍수터의 생태적 연결성을 조사하고 평가하는 것이 필요하다. 특히 하도와 홍수터에서 어류 군집의 특성을 파악함으로써 두 서식지의 연결성을 효과적으로 평가할 수 있다 (Miranda 2005, Zeug et al. 2005). 또한 본류 하도와 구하도의 연결성이 어류에 미치는 영향에 대하여 연구가 진행되고 있는데, 홍수터의 어류 산란장으로서의 역할과 본류와 홍수터의 어류 군집 차이에 대한 연구가 활발히 이루어지고 있다 (Rose and Baker 1983, Schiemer 2000, Zeug et al. 2005, Miyazono et al. 2010, Andrews et al. 2014).
만경강은 전라북도 완주군에서 발원하여 호남평야의 중심부를 지나 서해로 흘러드는 유역면적 1,527 km2, 유로 연장 약 77 km인 하천이다 (K-water 2007). 중하류의 평야지역을 지나는 이 하천은 과거에 사행이 심하였으나 정비사업으로 직강화되었다. 직강화로 인하여 사행하도 구간이 절단 (cutoff)되어 제내지에 약 7개의 구하도 구간이 형성되어 있다 (Hong et al. 2012). 이러한 구하도는 본류 하도와 단절되었으며 농경지, 주거지, 공사 등의 영향으로 어류 서식지가 교란 또는 파괴된 실정이다. 만경강에서 어류에 대한 연구는 주로 하천의 본류를 중심으로 전반적인 어류상 조사가 이루어 졌다 (Kim and Lee 1998, Lee et al. 2008). 그러나 만경강에 있는 구하도에서 어류 군집에 대한 정밀 조사는 이루어지지 않은 실정이다.
따라서 본 연구에서는 만경강에서 직강화 및 제방에 의하여 본류 하도와 단절된 대표적인 두 개의 구하도를 선정하여 1) 본류 하도와 구하도에서 어류 군집 구조의 특성을 비교하고, 2) 물리적 및 이화학적 환경 요인을 조사하여 본류 하도와 구하도에서 환경 차이를 분석하고, 구하도 서식 환경이 어류 군집 구조에 미치는 영향을 파악하여, 3) 이 결과를 바탕으로 구하도의 하천 환경을 개선할 수 있는 방안을 제시하고자 한다.
2. 연구방법
2.1 조사지 개황
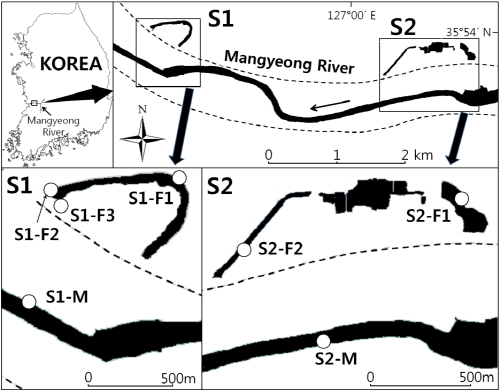
제내지 구하도와 본류 하도에서 어류 군집 구조의 차이를 비교하기 위하여 어류 채집과 환경요인 조사를 전라북도 익산시 만경강에서 2013년 9월에 실시하였다 (Fig. 1). 조사지점은 만경강에서 익산시 석탄동 S1 (35°54′N, 126°58′E)과 춘포면 용연리 S2 (35°53′N, 127°00′E)에 위치한 제내지의 격리된 구하도를 조사지로 선정하였다. 선정된 두 구하도에서 어류 서식 환경이 다를 것으로 예상되는 곳을 S1에서는 3개의 조사지점인 S1-F1 (35°54′15.6″N, 126° 58′31.4″E), S1-F2 (35°54′14.4″N, 126°58′12.6″E), S1-F3 (35°54′11.7″N, 126°58′14.8″E)을 S2에서는 2개의 조사지점 S2-F1 (35°54′05.1″N, 127°01′02.9″E) 과 S2-F2 (35°53′55.3″N, 127°00′24.2″E)을 선정하였다. 또한 두 구하도의 대조지로서 인근에 위치한 제외지 하도를 본류 조사지점 S1-M (35°53′58.7″N, 126°58′07.2″E)과 S2-M (35°53′38.3″N, 127°00′16.2″E)으로 선정하였다. 익산시 석탄동 (S1)에 위치한 구하도는 길이가 약 900 m이며 개방수면이 잘 발달되어서 양호한 어류서식처를 가진 구하도였다. 춘포면 용연리 (S2)에 위치한 구하도는 길이가 약 1.4 km이었고 일부 구간은 식생에 의해 개방수면이 줄어든 상태였다. 특히 본 조사지점은 매립공사가 진행되어서 구하도 내에서 일부 하도가 단절된 상황이었다. 두 조사지점 모두 주변은 주거지 및 농경지가 위치하였다. 본류 조사지점은 만경강의 본류로써 수로 폭이 넓고 하안에 식생이 발달하지 않았다. 물가에서 제방까지 주로 농경지가 발달한 상황이었다.
|
Fig. 1. Map showing the study area in the Mangyeong River, Korea. The opened circles indicate study sites. Dotted lines indicate river levee. |
2.2 어류 채집
선정된 조사지점에서 어류를 정량 채집하기 위하여 조사 시 투망 (망목 7×7 mm)을 15회 투척, 족대 (망목 4×4 mm) 30분 사용하여 채집하였다. 채집된 어류는 현장에서 동정하고 계수한 후에 방류하였다. 어류의 동정은 Kim and Park (2002)과 Kim et al. (2005)을 사용하였고 분류체계는 Nelson (2006)을 참조하였다.
2.3 환경 조사
하천 환경 조사는 미국 환경청의 하천 표준 조사법 (Lazorchak et al. 1998, Barbour et al. 1999)을 참조하여 총 9개의 환경 요인을 측정하였다. 하천의 물리 환경은 각 조사지점에서 임의로 20개를 선정하여 유속, 수심, 하상구조, 하도 횡단 폭 및 식생 피도를 측정하였다. 유속은 유속계 (Flowatch, JDC, Electronic SA, Switzerland)로 수심의 60% 지점에서 측정하였고, 수심은 접자를 사용하여 측정하였다. 측정된 유속과 수심은 평균으로 나타내었다. 하상구조는 Cummins (1962)의 방법을 참조하여 진흙 (mud, < 0.1 mm), 모래 (sand, 0.1 - 2 mm), 잔자갈 (gravel, 2 - 16 mm), 왕자갈 (pebble, 16 - 64 mm), 잔돌 (cobble, 64 - 256 mm), 호박돌 (boulder, > 256 mm)로 구분하였고 50 ×50 cm 방형구를 설치해서 방형구 내에서 가장 우세한 하상 입자를 기록하였다. 조사지점의 하상구조는 기록된 하상 입자의 상대빈도로 표시하였다. 각 지점에서 하도의 횡단 폭은 거리측정기를 사용하여 측정하여 평균을 구하였다. 조사지점 저수로의 식생 피도는 식생형을 구분하지 않고 육안으로 측정한 후 평균을 구하였다. 하천수의 이화학적 특성은 휴대용 수질측정기 (YSI 556 MPS, YSI Inc., USA)를 사용하여 수온, pH, 용존산소, 전기전도도를 측정하였다.
2.4 자료 분석
하천 환경에 따른 조사지점 사이의 관계를 분석하기 위하여 R 환경 (R Development Core Team 2013)에서 Vegan package (Oksanen 2013)의 ‘rda’ 함수를 사용하여 주요인분석 (principal component analysis, PCA)을 실시하였다. 입력 자료는 조사지점별 9개의 환경요인으로 구성하였다.
어류 군집 구조에 따른 조사지점 사이의 관계를 분석하기 위하여 R 환경의 Vegan package (Oksanen 2013)에서 ‘hclust’ 함수를 사용하여 계층적 군집분석 (hierarchical cluster analysis)을 실시하였다. 채집된 어종별 총 개체수(S)를 log(S+1)로 전환하여 입력 자료로 사용하였으며, 거리의 산출은 Bray-Curtis (Bray and Curtis 1957)의 방식을 적용하였다. 군집분석에서 산출된 높이 수준 (height level)을 기준으로 각 조사지점간 유사거리를 비가중산술결합 (unweighted pair group method using arithmetic algorithm, UPGMA) 방식 (Sneath and Sokal 1973)으로 분류하였다.
어류 군집 구조와 조사지점별 관계를 서열법 (ordi-nation)으로 분석하기 위하여 R 환경의 Vegan package (Oksanen 2013)에서 ‘metaMDS’ 함수를 사용하여 비모수다차원척도법 (non-metric multidimensional scaling, NMDS) (Kruscal 1964)을 적용하였다. 분석용 입력 자료는 종별 개체수를 log(S+1)로 전환하여 사용하였다. 조사지점 사이 거리는 Bray-Curits (Bray and Curtis 1957) 방식으로 산출하였다. 조사지 환경 요인과 2차원으로 도식화 된 NMDS결과의 상관관계를 분석하기 위하여 함수 Vegan package (Oksanen 2013)의 함수 ‘envfit’를 사용하여 상관계수와 유의수준을 산출하였다.
3. 결 과
3.1 환경 특성
만경강 구하도와 본류 하도의 모든 조사지점에서 측정된 수온과 pH는 유사하였다 (Table 1). 조사지점 S1-F3은 구하도의 작은 수로로서 수로 폭이 평균 3 m로 좁고 수심이 평균 23 cm로 얕았다. 하상은 진흙 (mud)과 왕자갈 (pebble)로 구성된 곳이었다. 조사지점 S1-F1와 S1-F2는 수로 폭이 평균 33 – 42 m로 S1-F3에 넓었고 수심 또한 평균 119 – 130 m로 깊었다. 하상재료는 대부분 진흙과 모래 (sand)로 구성되어있었다. 조사지점 S2-F1은 조사 당시 매립이 진행되고 있어서 교란 정도가 심했으며 수질이 조사지점 중 가장 좋지 않았는데, 특히 용존산소가 6.92 mg L-1로 가장 낮게 나타났다. 이곳에서 식생 피도가 평균 75.5 %로 가장 높았고 개방수면이 드물었다. 반면 조사지점 S2-F2는 S2-F1에 비해 수질이 양호하였으며 수심 평균 74 cm로 얕았고 식생 피도 평균 42%로 낮게 나타났다. 만경강 본류 조사지점은 구하도 조사지점에 비해 수로 폭이 평균 82 – 88 m로 넓었으며 식생의 피도가 낮게 나타났다. 이화학적 요인 중 전기전도도가 구하도 조사지점에 비해 높게 나타났다. 조사지점 S1-M은 수심이 평균 82 cm로 깊었고 물 흐름이 정체된 수역이었다. 하상은 주로 진흙과 모래이었다. 또 다른 본류 조사지점인 S2-M은 다리 주변으로 수심이 평균 38 cm로 얕았고 유수구간이 형성되어 있어서 유속이 평균 0.04 m s-1로 나타났다. 하상재료는 인공적인 거석이 있어서 잔돌 (cobble)과 호박돌 (boulder)의 비율이 높았다.
3.2 환경과 조사지점의 관계
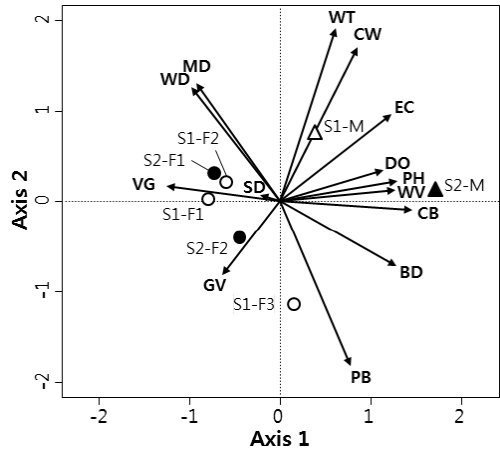
만경강의 조사지점별 환경요인 자료를 이용한 주요인분석 (PCA)에서 1축과 2축의 고유값은 각각 7.11과 3.29으로서 두 축이 전체 변이의 74.4%를 설명하였다 (Fig. 2). 환경 요인에 따른 조사지점의 배열을 살펴보면, 조사지점 S1-F1, S1-F2, S2-F1, S2-F2가 제 1축에서 왼쪽에 배열되었고, 조사지점 S1-F3는 제 2축에서 아래쪽에 배열되었다. 조사지점 S2-M이 제 1축에서 가장 오른쪽에 배열되었으며 조사지점 S1-M은 오른쪽 위에 배열되었다.
주요인분석 결과에서 환경 요인의 배열을 살펴보면, 식생 피도 (VG)가 왼쪽 가운데에 위치하였고, 수심 (WD)과 진흙 (MD)이 왼쪽 위에 배열되었다. 잔자갈 (gravel) (GV)은 왼쪽 아래에 배열되었다. 용존산소 (DO), pH (PH), 유속 (WV), 잔돌 (CB)은 오른쪽 중앙에 배열되었고, 수온 (WT), 하도 폭 (CW), 전기전도도 (EC)는 오른쪽 위에 위치하였다. 왕자갈 (PB)와 호박돌 (BD)은 오른쪽 아래에 배열되었다.
조사지점과 환경요인의 관계를 살펴보면, 구하도 조사지점 중 S1-F1, S1-F2 및 S2-F1은 식생 피도가 높고 수심이 깊으며 바닥이 진흙이 깔린 곳이었으며, 또 다른 구하도 조사지점 S2-F2와 S1-F3는 식생 피도가 높지만 하상이 잔자갈과 왕자갈로 구성되고 수심이 얕은 곳이었다. 반면에 본류 조사지점은 식생이 드물고 용존산소, pH, 전기전도도가 높고 하상이 잔돌로 이루어진 곳이었다. 특히 본류인 조사지점 S1-M은 하도 폭이 넓고 수온이 높았다.
3.3 어류상
만경강 전체 조사지점에서 채집된 어류는 총 258개체이며 10과 18종이었다 (Table 2). 과별로는 잉어과 Cyprinidae가 8종 (44%)으로 가장 많았으며, 그 다음은 망둥어과 Gobiidae가 2종 (11%)을 차지하였다. 만경강에서 우점종은 대륙송사리 Oryzias sinensis (26%) 였으나 주로 조사지점 S1-F3에 집중적으로 서식하고 있었다. 떡붕어 Carassius cuvieri (24%)는 아우점종이었으며 주로 구하도에 넓게 분포하였다. 다음으로는 붕어 C. auratus, 끄리 Opsariichthys uncirostris amurensis, 밀어 Rhinogobius brunneus, 좀구굴치 Micropercops swinhonis 순이었다.
구하도 조사지점인 S1-F1, S1-F2, S2-F2에서는 6종이 출현하였고 떡붕어가 우점종이었다. 이들 조사지점은 구하도의 중심 구간으로 물이 정체되고 수심이 깊고 하안에 식생이 잘 발달하였다. 특히 조사지점 S2-F2는 좀구굴치가 집중적으로 분포하고 있었다. 조사지점 S2-F1은 구하도 중에서 서식 환경이 가장 좋지 못한 곳으로 3종이 출현하였으며 특히 버들붕어Macropodus ocellatus는 본 조사지점에서만 출현하였다. 구하도의 작은 연결수로인 조사지점 S1-F3에서는 8종이 출현하였다. 대륙송사리가 우점하였으며 왜몰개 Aphyocypris chinensis, 얼록동사리 Odon-tobutis interrupta는 본 조사지점에서만 출현하였다. 만경강 본류 조사지점인 S1-M은 5종이 출현하였는데, 끄리가 우점하였으며 밀어와 모래무지 Pseudogobio esocinus가 우세하였다. 조사지점 St2-M은 5종이 출현하였으며, 끄리가 우점하는 것으로 나타났다. 피라미 Zacco platypus와 쏘가리 Siniperca scherzeri는 본 조사지점에서만 출현하였다.
3.4 어류 군집 구조
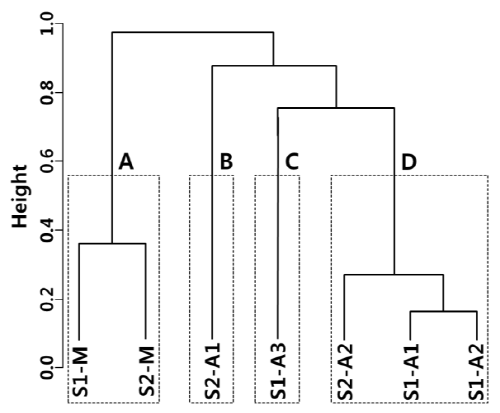
만경강에서 어류 종별 수도 자료를 사용한 계층적 군집분석에서 높이 수준 (height level) 0.58을 기준으로 집단 A (S1-M, S2-M), 집단 B (S2-F1), 집단 C (S1-F3), 집단 D (S1-F1, S1-F2, S2-F2)의 4개 집단으로 구분되었다 (Fig. 3). 집단 B, C 및 D는 구하도 조사지점으로서 서로 가까이 묶였으며 집단 A는 본류 조사지점으로 다른 집단과 멀리 구분되었다.
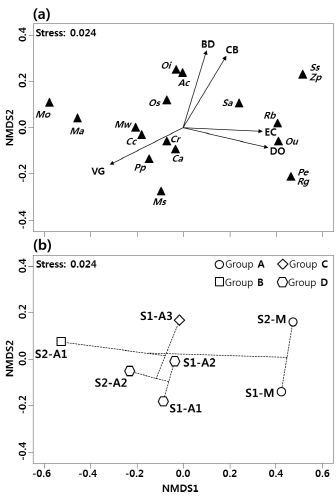
조사지점별 어류 종별 수도 자료를 비모수다차원척도법 (NMDS)에 의하여 2차원으로 분석하면 스트레스 (stress) 함수 값이 0.024로서 분석 결과의 설명력이 높았다 (Kruskal 1964) (Fig. 4). 비모수다차원척도법 결과에 계층적 군집 분석 결과를 투영하면, 구하도 조사지점 중 집단 D는 중앙 아래에 배열되었으며 집단 C는 중앙 위에, 집단 B는 왼쪽에 배열되었다. 만경강 본류 조사지점인 집단 A는 오른쪽에 배열되었다. NMDS 결과에서 종의 배열을 보면 버들붕어 (Mo)와 미꾸리 (Ma)는 수질이 좋지 못한 조사지점 S2-F1에 가까이 배열되었고, 왜몰개 (Ac), 대륙송사리 (Os), 얼록동사리 (Oi)는 조사지점 S1-F3 주변에 배열되었다. 구하도에 주로 서식하는 붕어 (Ca), 떡붕어 (Cc), 참붕어 Pseudorasbora parva (Pp), 잉어 Cyprinus carpio (Cr)는 조사지점 S1-F1, S1-F2 및 S2-F2에 주변으로 배열되었다. 또한 배스 Micro-pterus salmoides (Ms)는 유일하게 조사지점 S1-F1에 가까이 위치하였다. 좀구굴치 (Mw)는 조사지점 S2-F2에 가까이 배열되었다. 밀어 (Rb), 끄리 (Ou), 메기 (Sa)는 본류인 조사지점 S1-M, S2-M 주변에 위치하였으며 피라미 (Zp)와 쏘가리 (Ss)는 조사지점 S2-M에 가까이 배열되었다. 모래무지 (Pe)와 갈문망둑 Rhinogobius giurinus (Rg)은 조사지점 S1-M 주변으로 배열되었다.
3.5 어류 군집 구조와 환경과의 관계
만경강에서 비모수다차원척도법의 결과와 유의한 상관관계를 가지는 환경 요인은 용존산소 (DO) (r² = 0.97, P = 0.001, 5,000번 치환에 기초), 전기전도도 (COND) (r² = 0.82, P = 0.035), 잔돌 (CB) (r² = 0.91, P = 0.007), 호박돌 (BD) (r² = 0.84, P = 0.023). 식생 피도 (VG) (r² = 0.88, P = 0.014) 이었다 (Table 3). 이중에 잔돌과 호박돌은 NMDS 결과에서 위쪽으로 양의 상관관계를 보이고 있었고 집단 C와 가까이 배열된 왜몰개 (Ac), 얼록동사리 (Oi)가 위쪽 끝에 위치하였다 (Fig. 4a). 전기전도도와 용존산소는 집단 A 주변에 배열된 모래무지 (Pe), 피라미 (Zp), 끄리 (Ou), 메기 (Sa), 쏘가리 (Ss), 갈문망둑 (Rg), 밀어 (Rb)가 위치한 방향으로 갈수록 그 값이 높았다. 식생 피도는 구하도 조사지점이 배열된 왼쪽으로 갈수록 그 값이 높았고 그 끝 방향으로 붕어 (Ca), 떡붕어 (Cc), 잉어 (Cr), 참붕어 (Pp), 미꾸리 (Ma), 배스 (Ms), 좀구굴치 (Mw), 버들붕어 (Mo)가 위치하였다.
4. 고 찰
만경강에서 직강화에 의하여 단절된 두 개의 구하도와 본류에서 어류 군집 구조를 비교하였는데, 구하도와 본류 하도의 환경이 큰 차이를 보이고, 이에 따라서 두 곳의 어류의 생물상과 군집 구조가 큰 차이가 있음을 확인하였다. 구하도의 조사지점 S1-F1, S1-F2, S2-F2는 물이 정체되어 있고 물가에 식생이 발달하였으며 하상 재질이 진흙과 모래로 구성되어 있었다 (Table 1). 이들 조사지점은 다른 구하도 조사지점에 비해 정수성 어류가 서식하기에 양호한 환경이었다. 이곳에서는 주로 붕어, 떡붕어, 잉어, 참붕어 등이 서식하였다. 또 다른 구하도 조사지점인 S2-F2는 앞서 설명한 조사지점 S1-F1 및 S1-F2와 비슷한 환경이었으나 수심이 더 얕고 다양한 하상 재질이 나타났다. 이곳에서는 좀구굴치와 대륙송사리가 서식하였는데, 이 두 종은 대체로 수심이 얕고 식생이 잘 발달한 농수로나 소류지에 서식하는 어류이다 (Kim and Kim 1997, Kim and Park 2002). 또 하나의 구하도 조사지점인 S1-F3은 구하도와 연결된 배수 갑문 사이의 수로로서 수심이 얕고 수로 폭이 가장 좁아서 농수로와 비슷한 환경이었다. 본 조사지점은 조사지점 중 가장 다양한 어류가 출현하였고, 대륙송사리가 가장 우세하게 출현하였고 농수로에 주로 서식하는 왜몰개, 좀구굴치가 출현하였다. 반면 조사지점 S2-F1은 매립공사가 진행되었고 식생으로 인하여 개방수면이 줄어들었기 때문에 교란이 가장 심각하여 가장 낮은 어류 종다양성을 보였고 수질에 대한 내성이 강한 버들붕어가 본 조사지점에서만 출현하였다. 만경강에서 수행된 습지형 구하도의 기능 평가에서도 이곳이 가장 낮은 평가를 받았다 (Hong et al. 2012).
만경강 본류 조사지점인 S1-M과 S2-M은 구하도 조사지점과는 환경이 다르고 어류 군집 구조에서도 차이가 있었다. 본류에서는 수로 폭이 넓고 식생이 적게 분포하는 반면 용존산소와 전기전도도가 높고 하상재질이 다양하였다. 이곳에서는 구하도와 달리 끄리와 밀어가 우세하였다. 또한 모래의 비율이 높은 조사지점 S1-M에서는 모래무지가 분포하였으며 유수 구간이 발달한 조사지점 S2-M에서는 피라미, 쏘가리 등이 출현하였다.
하천에서 홍수터가 본류 하도와 단절되어 홍수터의 정상적인 생태적 기능이 상실될 경우 본류와 홍수터에 서식하는 어류는 악영향을 받게 된다 (Schiemer and Spindler 1989). 이러한 영향은 산란을 위하여 본류에서 홍수터로 이동하는 특이한 생활사를 가지는 어류뿐만 아니라 하천 본류에서만 서식하는 일반 어류 또한 영향을 받는다 (Andrews et al. 2014). 대부분의 어류 종은 물 흐름이 느린 정수 구간에서 산란하기 때문에 홍수터는 이들 치어 생산성에 중요한 기능을 한다 (Turner et al. 1994). 이러한 이유는 홍수터가 본류와 달리 물이 정체되고 식생이 발달하여 치어의 피난처와 섭식장소를 제공하기 때문이다 (Dewey and Jennings 1992). 만경강에서 직강화 사업은 홍수터를 본류로부터 격리시켰을 뿐만 아니라 기존에 있던 하도를 절단함으로써 제내지에 구하도를 형성시켰다. 이러한 결과로 격리된 홍수터와 구하도를 포함하여 전체 하천 환경에 큰 변화가 일어난다 (Toth et al. 1995). 실제로 만경강에서는 제내지 구하도에서는 퇴적으로 하도 수심이 얕아지고 정체수역이 형성됨에 따라서 식생이 번무하여 개방수면이 좁아지고 수질이 악화되는 환경변화가 나타났다. 또한 본류에서는 복단면으로 정비된 저수로 하상이 점차 저하하여 제내지 구하도의 하상보다 크게 낮아지는 것으로 나타났다. 본류 하상 저하는 구하도와 단차를 더욱 크게하여 구하도와 본류의 수리적 연결성을 더욱 악화시킬 것으로 예상된다. 이러한 환경 변화는 서식지 단순화를 초래하여 구하도뿐만 아니라 본류의 어류 종 다양성을 감소시키는 결과를 가져온다 (Schiemer 2000, Miyazono et al. 2010). 또한 장기적으로 격리된 만경강의 홍수터와 구하도는 본류와 상이한 환경을 가지게 되어서 어류 군집의 특성이 크게 차이가 나는 것으로 생각된다 (Erskine et al. 2005, Andrews et al. 2014). 만경강의 단절된 제내지 구하도에서는 현재 본류에서 서식하는 유수성 어류의 치어가 나타나고 있지 않기 때문에 수리적 연결성이 단절되어 이곳이 본류 어류의 산란처 및 은신처의 기능을 수행하지 못하는 것으로 판단된다.
5. 결 론
본 연구에서 만경강에서 구하도와 본류는 뚜렷한 환경 차이에 따라 어류 군집 구조의 특성이 달랐으며 구하도 내에서도 환경 차이에 의해 어류 군집 또한 차이가 있음을 알 수 있었다. 수심이 깊고 하상이 진흙으로 이루어 졌으며 식생 피도가 높은 구하도에서는 호소에 서식하는 어류가 출현하였고 수심이 얕고 환경과 하상이 왕자갈로 이루어진 구하도에서는 농수로에 서식하는 어류가 주로 출현하였다. 반면 수심이 깊고 식생 피도가 낮으며 다양한 하상구조로 이루어진 본류는 주로 대형 하천에서 서식하는 어류가 출현하였다. 이러한 결과는 다른 연구에서도 나타나는 어류 군집의 교란으로서 하천에서 횡적 연결성을 확보하는 필요성을 보여주고 있다 (Andrews et al. 2014). 또한 본류와 격리된 구하도의 지속적인 관리가 유지되지 않을 경우 수질악화, 토사퇴적 및 식생침입 등에 의해 어류 서식처와 어류 군집에 악영향을 줄 것으로 예상된다 (Miranda et al. 2001, Miyazono et al. 2010). 따라서 구하도의 지속적인 관리뿐만 아니라 본류와의 연결성 확보가 요구된다. 특히 본류와 구하도의 연결성이 확보될 경우 홍수와 같은 본류에서의 수리적 역동성 (hydraulic dynamics)이 구하도에 영향을 주게 되며 (Tockner et al. 1999), 이에 따라 서식지의 구조적 다양성, 물질 순환, 어류의 치어 생산성 등이 증가할 것으로 예상된다 (Rose and Baker 1983, Kwak 1988, Kingsford 2000, Miyazono et al. 2010).
따라서 본 연구 결과를 바탕으로 만경강에서 구하도의 환경을 개선하기 위한 방안을 다음과 같이 제안하고자 한다. 만경강에서 홍수터와 단절된 구하도와 본류의 수리적 연결성을 확보할 필요가 있다. 구하도의 횡적 연결성을 확보하기 위하여 제방을 후퇴하여 구하도를 제외지 공간으로 연결하는 것이 최선의 방법으로 생각된다. 제방을 후퇴하기 어렵다면 적절한 수문의 설치와 운영을 통하여 수리적 연결성이 가능하도록 개선할 수 있을 것이다. 현재는 배수 갑문과 배수 펌프가 설치되어 있어서 본류의 물이 구하도로 유입되지 못하고, 평수위에서는 배수문에서 구하도와 본류 하도의 단차가 너무 켜서 어류의 이동이 불가능하고 홍수위에서는 배수 펌프에 의하여 구하도의 물이 일방적으로 본류로 배제되고 있는 상황이었다. 그러므로 어류 이동이 가능하도록 제방에 수문을 설치하고 운영하는 것이 필요하다고 생각된다.
